降水变化对杉木种群动态的影响1)
2020-04-10冯明纪小芳姜姜
冯明 纪小芳 姜姜
(南方现代林业协同创新中心,江苏省水土保持与生态修复重点实验室(南京林业大学),南京,210037)
在1950—2010年,中国南部、中南部地区的年降水量都呈现减少的趋势,严重影响树木的生长[1-2]。降水量的减少以及相关的干旱会导致树木的死亡,造成森林的碳汇降低和整个区域森林枯死[3-4]。土壤水分的亏缺还会对植物体内碳分配和植物形态产生影响,植物会通过复杂的地上、地下活动影响土壤碳库的变化[5]。降水量变化对群落结构和种间相互作用的影响,最终将对群落生物量的稳定性产生影响[6-7]。杉木(Cunninghamialanceolate(Lamb.) Hook.)作为一种速生针叶树种,在中国广泛栽培,栽培面积达到1.7×107hm2,分别占中国人工林面积的24%和世界人工林面积的6.1%[8]。因此,在全球气候变化的大背景下,了解杉木种群对降水模式变化的响应规律和机制,对于更好地维护杉木林的持续性和稳定性具有重要的意义。
野外控制性试验在探究降水对土壤碳循环、养分循环、土壤呼吸影响[9-10]等方面发挥了重要作用,但是很难准确地判断降水变化对杉木种群结构的影响。由于树林木的寿命较长,且树木的种群动态、发展规律只有在很大的时空尺度下才会呈现。如果采用野外试验,将耗费大量的人力、财力、和物力[11],并且温度、降水、光照、病虫等因素都会对杉木生态系统产生影响,在现有的试验条件下,很难将各个因素区分开,这将对理解了解降水对杉木种群的影响产生巨大的阻碍[11-12]。不同条件下相互作用也可能产生相类似的结果,同一条件下也可能产生的结果差别很大[13]。因此,在面对复杂的生态学问题时,模型将帮助研究和理解某一个现象的形成过程和机理[14]。
本研究利用一个基于个体模型的森林动态生长模拟器[15],研究杉木种群动态及其对降水变化的响应,该模拟器采用了必要的降水、水分动态作用的模型。通过搜集参考文献数据及实地调查数据,对模型生长参数合理化设置。在此基础上,调整降水模式模拟杉木生长动态。相对前人的研究[16-17],用该物种生物量或个体数量的标准差和平均值的比值表征种群数量的时间稳定性,本研究对种群个体数量的方差和均值作线性回归(lnV=b×ln(M)+a),即Taylor幂法则中的b值表征杉木种群数量的时间稳定性[18]。Taylor描述了种群数量的方差与均值的关系,在昆虫、细菌等方面都得到验证[19]。在生态学中b值也常作为种群稳定性表征值[20]。
本研究将杉木作为研究对象,在中国和世界范围内,杉木都是一种很重要的生态、经济树种[21]。作为一种速生针叶树种,广泛栽培于中国的亚热带地区,分布于北纬20°~34°,东经100°~120°。杉木适应于年平均温度在15~23 ℃,年降水量800~2 000 mm的气候条件下生长。从十九世纪八十年代起,全球气温在以每10年0.065 ℃的速度上升,伴随着降水分配情况的改变,已经对杉木生长造成危害[22-23]。
本研究将探讨以下几个问题:杉木种群的生长是否遵循Taylor幂法则;年降水量的变化对杉木种群产生的影响;年降水量不变的情况下,季节性降水量的变化对杉木种群生长会产生的影响。期望通过阐明降水量的变化对森林生态系统的影响,为维护森林生态系统良性发展打下基础。
1 研究区概况
研究区位于江西省吉安市永丰县官山林场,地理坐标为北纬26°38′~27°32′,东经115°17′~115°56′,永丰县属亚热带季风气候区,四季分明,雨量充沛,日照充足,年均气温18.0 ℃,年均降水量1 627.3 mm,年无霜期279 d。在研究区内,设置20 m×20 m样方80个,作为数据来源和参照,现阶段仅用于初始林分设计的参照。
2 研究方法
2.1 杉木基本生长过程
基本生长过程主要包含输入、生长调节、输出三3个部分。输入对生长调节和输出都至关重要,决定着后续运行状态。生长调节连接着输入和输出两个部分,通过死亡、空间生长调节、降水生长调节和异速生长模型共同调节树木生长,并且实现树木之间的竞争调节。最终输出生长结构,以森林结构、光等形式与生长调节部分联系在一起(如图1)。
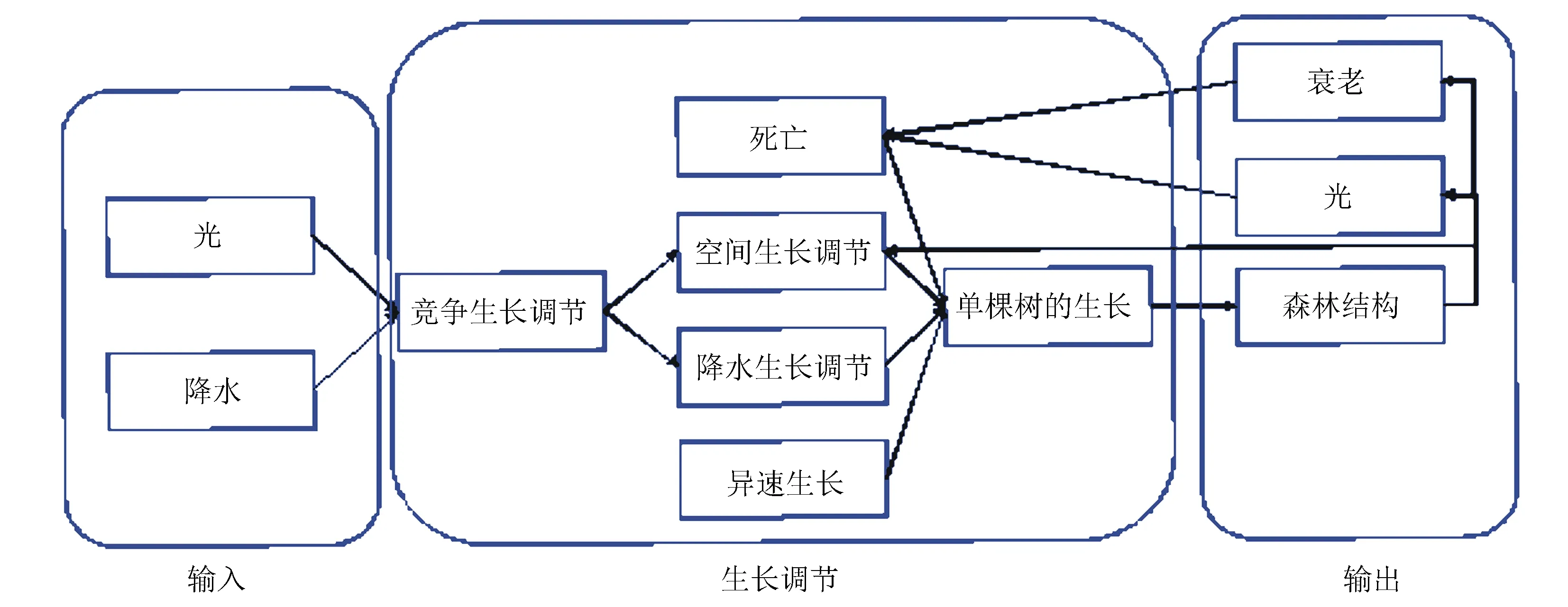
图1 杉木生长模型示意图
2.1.1 异速生长调节
树木在每个阶段表现出来的生理特性,不同生长速度对光和水的需要也不一样。因此,将树木的生长阶段划分为幼苗、小树、成年树,这样能更加真实的再现树木的生长过程。地径达到0.2 cm作为幼苗计算,树高达到1.35 m即为小树,胸径超过10 cm按照成年树处理。胸径(或地径)、树高和冠幅之间相互关联,因此,引入相关异速生长模型。分别采用标准胸径(地径)-树高生长模型、标准冠幅-胸径生长模型、标准冠幅深度-树高生长模型。
标准胸径(地径)-树高生长公式(适合树苗)h=0.1+30×(1-e(-a×d10));标准胸径(地径)-树高生长公式(适合小树和成年树的)h=1.35+(H1-1.35)(1-e(-b×DBH))。式中,h是树高;a为树高-地径参数;d10为树苗地上10 cm处的半径。H1为最大树高;b为树高-胸径参数;DBH为胸径。
2.1.2 生长光调节
为计算一棵树所获得的光,系统会模拟在每一棵树上安装一个鱼眼镜头,来确定被附近其它树遮挡的光照,计算从不同方向获得的光照,并计算出一棵树所获得的光。周围不同方向,树的大小不同,对目标树产生的影响也不同,对光照的计算模拟参考相关研究[24-25]。在本研究中,假设光对树木的生长不产生影响,仅影响树木的死亡,即当树木获得光照不足时,会出现一定死亡几率。
光照-死亡率调节公式为:m=m1×e-m2G。式中,m为一棵树死亡的可能性(概率);m1为一棵树在零生长时候的死亡率(2.5 a的死亡率);m2为与光照相关的死亡率[4](2.5 a的死亡率);G为胸径(地径)生长速率,5 a内生长累加。模型步长为5 a,计算5 a的死亡率公式:p′=1-(1-p)2。式中,p′为5 a的死亡率,p为2.5 a期间的死亡率。在研究季节性降水对森林的影响时,采用了另外一种计算方法。其公式为:m=1-exp(-(T×m1)×exp(-m2×G))。式中:T为一个步长所计算的时间长度;m为T时间内的死亡率;m1是在零生长速率时的死亡率;m2是与光照相关的死亡率[2];G为胸径(地径)生长速度,T周期内的生长量进行累加。
胸径-死亡率调节公式:
式中:ms为死亡概率;a和β为控制衰老上升幅度的相关系数;DBH为树木胸径;DBHs为树木开始衰老的最小胸径。
2.1.3 树木死亡及种子传播
每棵树都会随机死亡,在一定的时间和面积都会有一定数量的树木死亡。应用一个基于生长速度的死亡模型,以体现树木在自然状态下被淘汰的情况。应用衰老模型评判其衰老死亡概率。在现实中当树木没有到达一定的胸径时,很少会因为衰老而死亡,当超过一定胸径,因衰老而死亡的概率逐渐增大,因此,设置了胸径相关的死亡概率模型。种子传播要依赖于母树的位置和大小来决定产生种子的数量和位置,种子产生的位置通过威布尔概率分布模型来确定。
2.2 降水对生长的调节
2.2.1 年降水量对生长的调节
通过利用一个基于邻里竞争生长模型(NCI)来调控树木的生长速度[26],该模型可以选择多个影响因子进行组合,形成竞争关系进而限制树木的生长。为研究降水对树木生长的影响,排除养分等干扰因素,本研究选取了空间竞争系数、邻里胁迫系数和降水调节系数进行组合。对一棵树木的生长,首先设置了最大潜在生长量,邻里竞争生长模型公式如下:G=MG×CE×DE×PE。式中,G为胸径(地径)生长速率;MG为最大潜在胸径(地径)生长速率;CE为空间竞争系数,DE为邻里胁迫系数,PE为降水调节系数,各个系数值均处于0~1。
2.2.2 季节性降水对生长的调节
对生长部分的调节,同样采取空间竞争系数、邻里胁迫系数和降水调节系数因素组合。不同的是在降水调节上,采用季节性降水亏缺指数进行计算,该指数在年均气温和降水量的基础上进行计算,实现了水热循环过程。该指数为当前水分条件下潜在蒸发量与可利用水分条件下的实际蒸发量的差值,所以该指数可以解释实际水分能否满足供应需求量。年实际蒸发量为各月实际蒸发量之和,年潜在蒸发量为各月潜在蒸发量之和。季节性降水亏缺指数为潜在蒸发量与实际蒸发量的差值,该值将用于降水竞争指数的计算。
本研究中气象数据参考2017年江西省吉安县气象站数据,下载于中国气象数据网。
2.3 方案设计
本研究初始林分密度为20株/hm2,均为幼苗。样方大小设计为100 m×100 m,其中有25个20 m×20 m的统计单元。为探究不同年份降水量情况下的杉木的种群特性,及季节性降水变化对杉木的影响,设计年降水量为1 000~2 000 mm,梯度为100 mm,共11种年降水强度,同时对各季节的降水量进行调控。根据杉木的生长特性,将1年分为生长季和非生长季2个部分,生长季按5—6月、7—8月、9—10月等,分别对应杉木生长的初期(春季)、旺盛期(夏季)、末期(秋季),对照组的各月份降水量参照2016年江西省吉安县各月实际降水情况,处理组则对生长季降水量按比例进行增减(30%~-50%,梯度为10%),增减降水量均匀分配到非生长季月份[27],以保证年降水量恒定。
2.4 数据处理
模型在每一步运行结束后,会输出各个树木的位置、胸径(地径)、高度和冠幅等信息,由于数据量巨大,本研究采用MATLAB对树木坐标进行数据化处理。通过MATLAB将样方内每个20×20单元格内的树木数量进行统计,得到25个单元格内树木数量的均值(M)和方差(V),在时间序列下,共计得到120对均值和方差数据,对每个处理得到数据取对数,取森林平衡前的均值和方差数据做线性回归。
3 结果与分析
3.1 Taylor幂法则验证
由图2可知,通过对20组数据的分析表明,单元格内树木株数的方差的对数(lnV)与其均值的对数(lnM)均呈现良好的正相关关系(R2>0.95),得到线性回归方程lnV=b×(lnM)+a,本模型结果验证了Taylor幂法则,种群稳定性指数b值处于0~2,与其他领域相关研究的结果一致[28-29]。
3.2 年降水量对杉木种群的影响
由图3可知,年降水量对杉木的生长状态有显著影响,生长平衡时,杉木种群密度、胸径与单位林木断面积均呈现对称的抛物线型。在降水量低于最适降水量时,随着降水量的增加,杉木种群密度、胸径及单位林木断面积均呈现增长趋势,但增长速度减缓;在降水量高于最适降水量时,随着降水量的增加,这些指标都呈现下降趋势。随着降水量的增加,b值(图像的斜率)有显著下降的趋势。

图2 降水量和生长季降水移除时的树木lnM与lnV关系图

图3 杉木种群评价指标与年降水量关系
3.3 季节性降水对杉木种群的影响
由图4可知,季节性降水在一定变化范围内对杉木的生长有显著影响,长期而言,季节性降水增加时,对杉木种群密度、平均胸径、胸高断面积没有影响。当生长季降水减少30%以上时,生长季降水的变化对杉木种群密度、平均胸径、胸高断面积有显著影响。在减少20%以下的季节性降水量或增加季节性降水量时,杉木的种群密度、胸径及胸高断面积都处于较为稳定的状态;减少季节性降水量超过30%时,杉木种群密度、平均胸径、胸高断面积呈现显著下降趋势。相对于对照组,随着季节性降水量的减少或增加,b值都会增加。
4 结论与讨论
4.1 Taylor幂法则在林业的应用
本研究中,杉木种群方差的对数与均值的对数均存在线性关系。大量研究证实许多物种都遵循Taylor幂法则[30-35]。Taylor et al.[18,30-32]通过对昆虫的研究,表明昆虫的种群数量遵循Taylor幂法则;Ramsayer et al.[36]也证明细菌同样遵循Taylor幂法则;有关学者在理论上对Taylor幂法则进行了论证[29,37]。但是,相对于昆虫、鱼类等,森林的寿命较长,研究难度较大,Taylor幂法则在林业的应用较为少见。Cohen et al.[38]对橡树(Quercusspp)的研究证明了空间Taylor幂法则,在此基础上,本文通过对杉木林的研究,验证了时间Taylor幂法则,种群数量的方差和均值取对数后的比值b均处于0~2,与其他领域相关研究结果一致。

图4 杉木种群评价指标与生长季降水量变化的关系
4.2 年降水量与杉木种群稳定性
降水量的减少对土壤弱酸化有一定的影响,会减少森林土壤有机物质的累积以及土壤向树木的营养供给[39]。土壤水分与养分之间存在着长期或短期的互作过程,在降水多而潮湿的地区,成土母岩风化更为强烈,形成的土壤酸化趋势更明显,加速土壤中的腐殖质及粘土向下迁移,而矿质营养离子及其他可溶性化合物则淋溶至渗透到地下水中。湿润的土壤通过质流和扩散为根系提供更好的营养条件,但土壤缺水干燥时,这两个过程都会受到抑制[40]。研究表明,热带潮湿山地森林土壤碳库随降水量的增加而增加,土壤有效氮含量则随降水量增加而减小[41]。
降水量和土壤水分作为土壤生物活动的控制性因素,影响着土壤中的物质分解和养分循环[10]。降水量的减少常与高温相互关联,温度升高加速土壤有机质分解,这对微生物群落的稳定生存是不利的,最终将影响菌根真菌群落结构,进而影响其对保护幼苗成活、抵御病原菌伤害和共生体的营养吸收的促进作用。菌根真菌连接着植物和土壤,形成植物-土壤反馈调节机制,植物会为菌根真菌提供营养[42-43]。当降水量减少、而平衡被打破,真菌群落系统和植物种群的稳定都将受到威胁。在本研究中最适年降水量情况下,树木生长都能达到最佳状态,但是该降水条件下杉木林种群时间稳定性低于降水量高的条件下的杉木种群时间稳定性。在更高的降水条件下,土壤能向杉木提供更多的养分,真菌促进养分循环,两者共同促进了杉木种群的时间稳定性。在年降水量升高情况下,杉木种群数量的时间稳定性也会随之增大。
4.3 季节性降水与种群稳定性
季节性降水对森林的各个方面都会产生影响。在保持年降水量不变的条件下,增加生长季降水、减少非生长季降水量,能够有效增强树木的蒸腾作用,使得树木快速生长。但是,这种条件下真菌群落的多样性明显下降[44],杉木获得营养的途径受到一定的限制[43],因此,在这种水分条件下,生长季降水量即使增加了,树木的生长量不会受到太大的影响。然而,当生长季降水量减少时,树木的生长严重受到限制,极端干旱成为主要限制因素[45],植物可利用水分减少,森林生态系统处于恶化状态,地上生物量降低,杉木种群稳定性随着生长季降水量的减少而降低。本研究表明,当生长季降水量增加时,杉木林的种群稳定性也会下降。这种现象是由杉木的生理特性所决定的,杉木的生理特性决定了其生态位,当环境的改变到达一个临界点时,杉木就不能够更好适应,并且必然有一个能够更加适应当前环境的物种出现,在接近临界点之前,其种群稳定性会越来越低,最终这一种群将消失,被另一种群取代。由此表明,当生长季降水量增加或减少时,杉木的种群稳定性都会减弱,会向下一个种群演变。
