长期稻-稻-紫云英轮作对水稻种子内生细菌的影响及促生功能评价
2020-04-08张子强夏振远KyuKyuThin郭鹤宝何山文高菊生张晓霞
赵 霞,张子强,夏振远,Kyu Kyu Thin,郭鹤宝,何山文,高菊生,4*,张晓霞*
(1.中国农业科学院农业资源与农业区划研究所,北京 100081;2.北京城市学院生物医药学部,北京 100083;3.云南省烟草农业科学研究院,云南 昆明 650021;4.中国农业科学院衡阳红壤实验站,湖南 祁阳 426182)
种子的生理生态学和病理学相关研究表明,植物种子内富含微生物种群[1-2]。利用可培养方法,在水稻、玉米、大麦、苜蓿等25种植物中,发现有4个门131个属的种子内生细菌,其中Proteobacteria为最主要的类群,其它依次为Actinobacteria,Bacteroidetes和 Firmicutes[3]。在属水平,Bacillu,Pseudomonas,Paenibacillus,Micrococcus,Staphylococcus,Pantoea等类群最为常见。近年来高通量测序技术应用到种子微生物的研究中,使得种子中微生物多样性得到进一步的认识。16S rRNA高通量测序发现种子中的主要类群与可培养法一致,为Proteobacteria,Actinobacteria,Bacteroidetes和Firmicutes,同时还有一些新类群如Acidobacteria,Fusobacteria,Chlamydiae和 Gemmatimonadetes[4-7]。种子内生菌也显示出生物防治和植物促生的功能,如固氮、溶磷、分泌生长素(IAA)和嗜铁素以及ACC(1-氨基环丙烷-1-羧酸)脱氨酶活性等[8-10]。此外,种子内生菌对其宿主植物具有抗致病菌、抗动物摄食等作用[11],可有效提高种子萌发率、幼苗存活率、幼苗根长和苗高、生物量积累等[12]。当植物种子落入土壤后,在萌发过程中,还会通过特定机制招募特定微生物在其表面聚集,以帮助种子萌发,防止退化或被捕食[13]。种子微生境目前研究较少,近年来因其作为有益微生物的储库和载体的潜力而备受关注[14]。
绿肥作为一种传统的农业技术,应用历史悠久。在化肥大规模应用之前,是增加土壤养分供应的主要来源之一。我国是世界上种植绿肥面积最广的国家。据不完全统计,19世纪中期,全国绿肥种植面积有173万hm2,随后逐步增加。到20世纪80年代初期,种植面积稳定在1 000万hm2左右,但到80年代中后期,由于化肥的推广和使用,绿肥生产开始出现滑坡,到90年代初期种植面积剧减[15-16]。虽然化肥在一定程度上提高了作物的产量,但由于化学合成物质在农田的大量使用严重改变了土壤原有的微生物组成和生物地球化学循环[17-18]。给农业生产和环境带来了一系列的负面作用[19-21],成为进一步提高作物产量及质量的重要障碍。而种植绿肥不仅能为土壤提供丰富的养分,控制植物病虫害的发生[22-23],还可以促进土壤微生物的活动[24-26],具有显著的土壤增肥功能[27]。实验室前期工作已证实长期水稻-稻-绿肥轮作可以显著改变水稻根际和根内生微生物群落的组成结构[28-29]。本研究进一步探讨了绿肥(紫云英)轮作对水稻种子内生细菌微生物群的影响与功能。
1 材料与方法
1.1 样品采集
长期绿肥与水稻轮作试验田位于湖南省祁阳县农业部红壤生态环境重点田间监测试验站(26°45′42″N,111°52′32″E)。1982~ 2012年,进行稻-稻-冬闲(RR-WF)和稻-稻-紫云英(RR-MV)轮作。每个处理重复3次,随机区组排列,小区面积37.5 m2(2.5 m×15.0 m),各小区之间由60 cm水泥埂隔开。每季早稻和晚稻的使用总肥料(基肥+追肥)包括N(153.0 kg/hm2),P2O5(84.0 kg/hm2)和K2O(129.0 kg/hm2)。复合肥[N(84.0 kg/hm2),P2O5(84.0 kg/hm2) 和K2O(84.0 kg/hm2)]用作基肥,N(69.0 kg/hm2)和K2O(45.0 kg/hm2)用作追肥。移栽水稻前施用基肥,水稻移栽6~10 d后追肥。在晚稻收获前10~15 d,播种绿肥紫云英种子(播种量为7.5 kg/hm2)。种植绿肥的小区在次年早稻移栽前15 d将鲜草全部翻入各自小区内。每季除稻茬外,其他秸秆全部移走。冬种绿肥不再施肥。2013年10月,每块地随机选取3个点采集水稻种子样品,风干后,4℃冷藏保存。
1.2 水稻种子表面杀菌和细菌分离培养
根据Sun等[30]的方法对水稻种子进行表面杀菌:两个处理各选取1 g种子,先用无菌水冲洗一次,然后分别浸入70%酒精和新鲜NaClO溶液(2.5%可用Cl-)中各3和5 min,再浸入70%酒精30 s,最后用无菌水清洗5次。将种子轻轻在TSA平板上按压翻滚以检查表面杀菌效果。将表面杀菌后的样品在装有少量无菌石英砂的研钵中研磨至粉末,然后用无菌水稀释成10-1、10-2和10-3的悬浮液。3个梯度各吸取100 μL分别在TSA[31](Difico),1/4 R2A[32](Difico) 和 Ashby[33]无 氮培养基(蔗糖20 g,K2HPO40.1 g,KH2PO40.4 g,MgSO4·7H2O 0.2 g,NaCl 0.01 g,FeCl30.01 g,Na2MoO4·2H2O 0.002 g,1 L水)上进行涂布,每种平板3个重复。所有平板28℃下培养3 d。挑取每个处理的1/4 R2A平板上的所有菌落,并从其他平板上挑取大小和颜色不同的菌落进行划线纯化。所有纯化后的菌株通过冷冻干燥法在4℃储存。
1.3 系统发育分析
使用DNA提取试剂盒(Transgen,China)提取细菌DNA(具体步骤参照试剂盒说明)。根据Lane[34], 使 用 通 用 引 物 27F 和 1492R 进 行 16S rRNA基因的扩增,PCR产物送生物公司测序。测序结果使用EzTaxon进行序列比对[35];系统发育树由 MEGA 6.0 软件构建[36]。
1.4 检测分离菌株的植物促生功能
在Pikovskaya培养基上进行菌株溶磷活性的检测[37];在含有100 μg/mL L-色氨酸的TSB培养基接种细菌并培养48 h,参照Glickmann等[38]的方法检测悬浮液中IAA含量;参照Machuca等[39]的方法检测菌株是否产铁载体。
2 结果与分析
2.1 可培养种子内生菌的数量
本研究使用3种不同的培养基估测水稻种子内生细菌的数量。平板计数结果如图1所示:3种不同培养基的计数结果均表明长期绿肥轮作显著增加水稻种子内生细菌的数量;同时绿肥轮作系统收获的水稻种子内生菌对培养基有不同的偏好:长期绿肥轮作R-R-MV在TSA上观察到的内生细菌数量最高,达2.87×106CFU/g,而冬闲R-R-WF在1/4 R2A上检测的细菌数量最高,为2.33×106CFU/g,说明R-R-MV和R-R-WF的种子内生细菌间存在不同的代谢类型。

图1 梯度稀释涂布并计数的方法检测水稻种子内生细菌的数量
2.2 可培养内生细菌系统发育分析
据梯度稀释平板细菌生长情况,选择1/4 R2A平板上所有菌落,和其他两种培养基上不同菌落形态不同颜色的细菌进行分离培养。结果R-R-WF种子分离到42株内生细菌,R-R-MV种子分离到66株内生细菌,通过16S rRNA基因序列扩增并测序,约700 bp序列上传至Eztaxon(http://www.ezbiocloud.net/eztaxon)比对分析;序列提交GenBank并获得登录号,图2、3括号中的数字即为相应菌株的GenBank登录号;使用MEGA 6.0构建系统发育树(图2、3)。结果表明,42株R-R-WF内生菌分别属于9属12种,66株R-R-MV内生菌属于15属21种。
R-R-WF和R-R-MV内生菌的优势门均为γ-Proteobacteria,分别占60%和50%。α-Proteobacteria(24%)是R-R-WF的次要优势群,其次是Actinobacteria和Firmicutes。而Firmicutes(22%)是R-R-MV的次要优势门,其次是Actinobacteria(15%)和α-Proteobacteria(9%)。而β- Proteobacteria(5%)仅在R-R-MV中检测到。
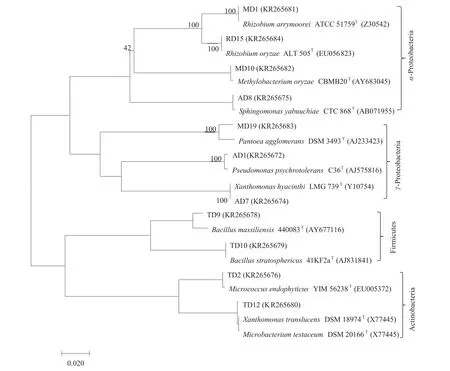
图2 R-R-WF水稻种子分离到的内生细菌16S rRNA序列的系统发育树
在属水平上,Pseudomonas(28.57%)和Xanthomonas(26.19%)是R-R-WF中的优势属,其次是Sphingomonas(16.67%)。 此外,在R-R-WF中还发现Bacillus,Rhizobium,Micrococcus,Methylobacterium,Pantoea和 Microbacterium。与R-R-WF相比,Pantoea(27.94%)是R-R-MV中的优势属,其次是Pseudomonas(14.71%),Paenibacillus(13.24%)。Paenibacillus,Curtobacterium,Acidovorax,Rothia,Acidovorax,Kocuria 和 Staphylococcus是R-R-MV中的特有类群(表1)。

图3 R-R-MV水稻种子分离到的内生细菌16S rRNA序列的系统发育树

表1 水稻种子内生细菌在属水平的相对丰度(R-R-WF:稻-稻-冬闲,R-R-MV:稻-稻-紫云英)(%)
2.3 种子内生菌植物促生特性检测
基于内生细菌的来源与分类地位,本研究选择31株菌检测植物促生特性(表2)。其中,14株( 属 于Bacillus,Curtobacterium,Kocuria,Microbacterium,Paenibacillus,Pantoea,Pseudomonas,Rhizobium,Rothia和Sphingomonas)可以分泌生长素IAA;10株(属于Bacillus,Curtobacterium,Microbacterium,Pseudomonas,Sphingomonas和 Xanthomonas)在CAS平板上的菌落周围产生橙色晕圈,表明这些菌株具有产铁载体的活性;9株可以溶解有机和无机磷酸盐(菌落周围可以观察到透明圈),测量菌落直径(n)和晕圈直径(z),计算z/n值,发现在无机磷酸盐培养基上,菌株 RZ27、TZ26和RZ4的z/n大于2.0,表明这些菌株具有强溶解无机磷的能力[40]。此外,8株菌可溶解有机磷酸盐,但不能溶解无机磷酸盐。TZ22、TD10和AD1具有本研究中所检测的植物促生的所有特征,但也有9株没有检测到任何促生作用。
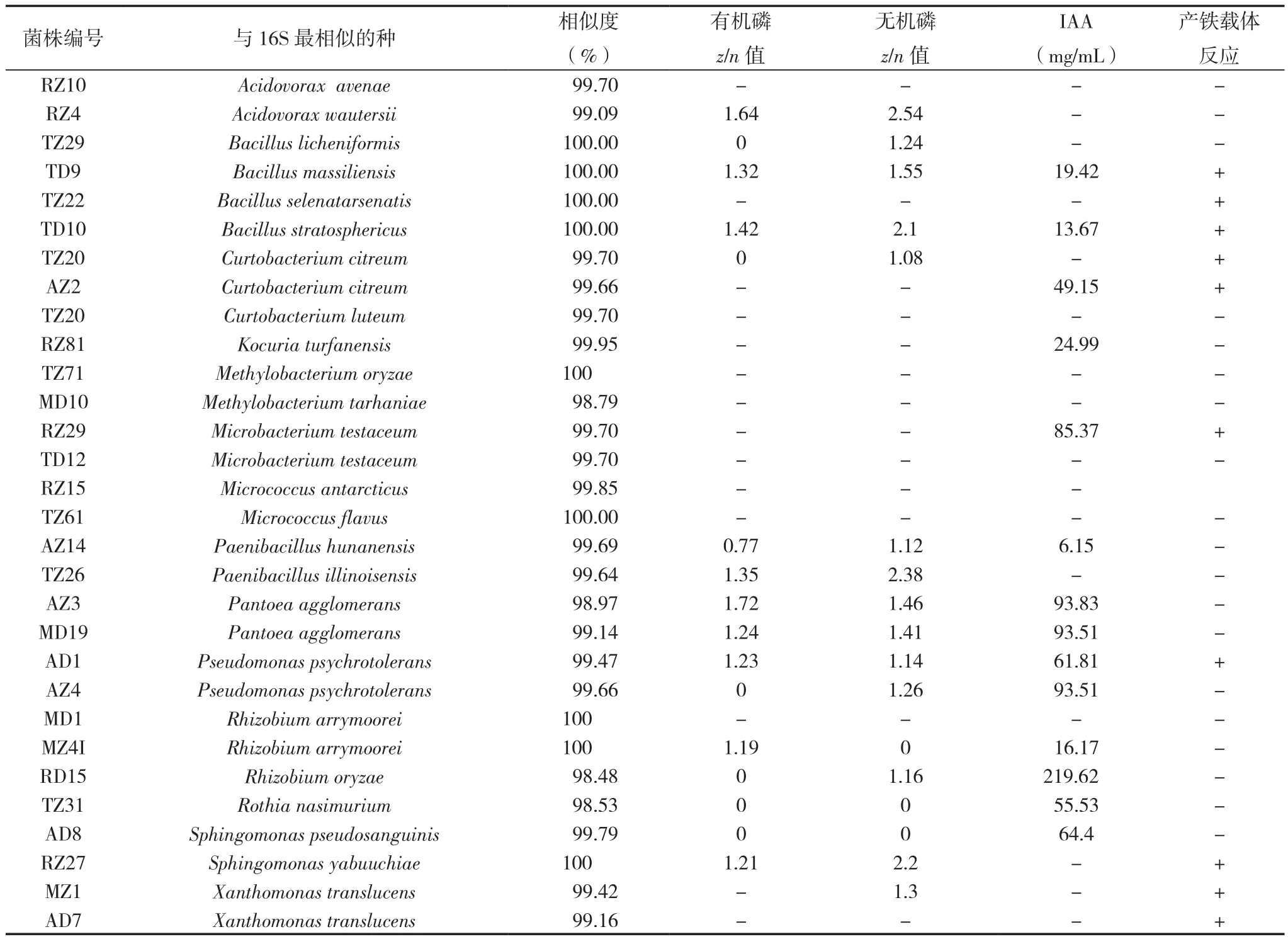
表2 水稻种子细菌的溶磷酸盐、IAA和产铁载体促生能力的检测
3 讨论
3.1 长期绿肥轮作下,水稻种子内生细菌变化情况及其与根内生细菌的关系
本试验前期研究已经发现长期紫云英绿肥轮作(R-R-MV)会显著改变水稻根内细菌的多样性和组成[28]。本研究通过传统分离培养方法证明长期紫云英绿肥轮作对水稻种子内生细菌的影响。结果表明内生细菌数量显著增加,多样性也明显增加,主要类群发生变化。可见长期的稻-稻-绿肥轮作不仅会改变水稻根内生细菌[28],也改变了种子内生菌的数量与多样性。与Zhang等[28]描述的根内生细菌相比,Pantoea,Pseudomonas,Xanthomonas和Sphingomonas是种子内生菌的主要类群,但在水稻根内丰度较低。Herbaspirillum和Cedecea是根内生细菌的优势属,但种子中并没有分离到。一些丰度较低的属(Methylobacterium,Curtobacterium,Rothia和Kocuria)仅在种子中分离到。Mano 等[41]根据大量关于水稻根和种子内生细菌的报道也得出类似的结论,水稻种子内的固有细菌较少,不同植物组织其菌群结构也不同。因此,本研究推测种子内生细菌受根内生菌和其他环境因素的影响。
3.2 水稻种子内生细菌的组成及其功能
Pseudomonas,Xanthomonas,Sphingomonas和Pantoea是本研究检测到的种子内生细菌优势属。此外,水稻种子中也存在非优势属,例如Paenibacillus,Curtobacterium,Acidovorax,Kocuria,Staphylococcus和 Rothi。Mano等[41]报道了水稻种子内生细菌主要包含Pantoea,Herbaspirillum,Metylobacterium,Klebsiella,Acidovorax,Bacillus,Curtobacterium,Micrococcus,Paenibacillus,Sphingomonas,Xanthomonas,Ochorbacterium,Pseudomonas等。过去基于16S rRNA Illumina测序发现Pantoea,Acinetobacter,Xanthomonas,Bacillus,Flavobacterium,Stenotrophomonas,Neorhizobium和 Pseudomonas是水稻种子中的共有优势属[7]。因此,本研究分离的细菌大多数是典型的种子内生细菌。
Pantoea是水稻种子内生细菌的主要种类之一,表明Pantoea和水稻之间有很强的亲和力。研究报道玉米种子内生细菌最丰富的也是Pantoea[42],Pantoea具有固氮、分泌植物激素的功能[43],本研究发现Pantoea具有产IAA、溶解无机和有机磷酸盐的能力。
Pseudomonas也是种子内生细菌的主要组成之一。本研究选择两株Pseudomonas来检测植物促生功能,发现AD1能够产IAA、铁载体,并可以溶解无机和有机磷酸盐(表2)。此外,Pseudomonas也是研究细菌与植物互作的模式菌[44],如Pseudomonas fluorescens可以通过分泌抗生素和竞争营养,使植物免受致病菌的侵害[45],并增加作物产量[46]。
本研究发现一些低丰度细菌对植物具有潜在的有益功能,如表2所示的Bacillus,Paenibacillus,Methylobacterium和Sphingomonas。这些菌株在不同的环境中被发现与植物有密切的关系。Methylobacterium具有很强的抗逆性,能够合成植物激素并刺激种子萌发[47]。Curtobacterium是一类典型的种子共生细菌,能够保护植物免受Erwinia的攻击[48-49]。Bacillus可溶解无机或有机磷酸盐,分泌IAA并产生抗真菌代谢物以抑制致病菌[50-51]。
4 结论
本研究的主要结论如下:(1)长期绿肥轮作改变了水稻种子内生细菌的群落结构和数量;(2)种子选择并积累了一些特定的细菌,如Pseudomonas,Xanthomonas,Sphingomonas和 Pantoea;(3) 大 多数分离菌株具有植物促生功能,表明它们与植物有密切关系。
