人工增加UV-B辐照改善温室番茄品质
2020-04-07郭建军王晓芳孙玉楚李景原
郭建军 王晓芳 孙玉楚 李景原


摘 要: 利用小型研究型温室,人工增加UV-B(紫外线B),旨在探讨增加UV-B辐射对改善温室番茄果实活性物质含量、提高番茄营养健康性能和商品品质的可行性,为温室生产提供参考。试验结果表明,适度增加UV-B辐照后,番茄节间长度缩短,株高降低。但植株结果节数增加,番茄果实数量增多,果实体积略有减小。UV-B辐射诱导花青素、黄酮类化合物等次生代谢物质的积累,刺激果实抗氧化能力和抗氧化活性的提升。因此,温室适量增加UV-B辐射,能提高番茄的营养物质,改善温室番茄果实品质。
关键词: 番茄; 抗氧化剂; 代谢调节; 营养价值; 酚类化合物; 紫外线辐照
中图分类号:S641.2 文献标志码:A 文章编号:1673-2871(2020)02-031-06
Effect of UV-B on the tomato quality in greenhouse
GUO Jianjun1, WANG Xiaofang2, SUN Yuchu2, LI Jingyuan2
(1. Zhengzhou Vocational College of Finance and Taxation, Zhengzhou 450048, Henan, China; 2. College of Life Science, Henan Normal University, Xinxiang 453007, Henan, China)
Abstract: In order to explore the feasibility of increasing UV-B radiation on improving the content of active substances in tomato fruits and the commodity of tomato in the greenhouse, and to provide reference for greenhouse production, the artificially increasing UV-B radiation in greenhouse was studied in this research. The results showed that the internode length and plant height decreased after increasing UV-B irradiation, however, the number of nodes increased, the number of tomato fruits increased, and the fruit volume decreased slightly. UV-B radiation could induce the accumulation of anthocyanins, flavonoids and other secondary metabolites, which stimulated the antioxidant capacity and antioxidant activity of fruits. Above all, increasing UV-B radiation in greenhouse could improve the nutritional quality and fruit quality of tomato.
Key words: Tomato; Antioxidants; Metabolism modulation; Nutritional value; Phenolic compounds; Irradiation of UV-B
新鮮番茄果实富含叶酸、维生素C、番茄红素和类黄酮等有益于人类健康的生物活性物质[1],番茄是大众消费的果蔬类之一,可以露天种植,也可在园艺设施(如温室)中种植以满足淡季需求。温室生产虽然弥补了淡季人们对新鲜番茄的需求,但露地大田生产的番茄果实更受消费者青睐,消费者普遍认为露地番茄果实的感官和营养质更佳[2],因此,提高温室种植番茄果实的感官和营养品质,对温室番茄生产尤其重要。
现代温室通常使用比玻璃便宜的聚碳酸酯,聚碳酸酯只能透过波长大于400 nm的光波,因此,现代温室种植的番茄紫外线辐射很弱,甚至没紫外线辐射[3]。已有研究证明,虽然紫外线辐射会对植物造成损伤,但也能诱导植物产生花青素、多酚类次生活性物质以提高植物对紫外线辐射的防御能力[4-5]。研究表明,隐花色素(Cry1和Cry2)、UVR8、植物色素和光调节素等光感受器在特定代谢途径分子调控中起着重要作用[6-9]。为了应对紫外线照射引起的活性氧产生和氧化应激的增加,光感受器会启动保护机制,导致酚类化合物和酶等抗氧化剂的增加[10]。酚类化合物在植物光保护方面具有重要的作用,紫外光可激活苯丙氨酸氨解酶、查尔酮合成酶和黄酮醇合成酶[11]。这些刺激最终会增加果实中类黄酮和类胡萝卜素等不同多酚的含量和比例,而这些多酚类抗氧化剂含量的增加有益于消费者的营养健康。同时,通过类黄酮和类胡萝卜素途径合成了多种挥发性有机化合物,这些物质对水果的风味和口感有着重要贡献[12]。笔者旨在通过人工增加适量的UV-B(紫外线B)辐射提高温室番茄的营养品质,改善温室番茄的商品品质。
1 材料和方法
1.1 植物生长条件和UV-B处理
番茄种子‘凯特莉由新乡市大自然农业技术开发有限公司培育,河南师范大学国家生命科学实验示范中心繁育保存。试验于2018年4—10月在河南师范大学温室中进行。将种子点播于V泥炭∶V珍珠岩=2∶1的营养钵(50 cm)中,然后种植在小型温室(5.0 m×9.0 m×3.6 m)的光照培养架上,周期为16/8 h(光/暗),光照强度为200 μmol·m?2·s-1 ,温度(23±2) ℃,相对湿度(45±5)%,每周灌溉2次,pH为5.70。90 d出现第1花序,第1花序出现10 d后,每组10株,分5组进行增加UV-B辐射试验。
采用完全随机设计,将番茄随机分成5组。第1组不加UV-B灯(对照组)。第2~第5组植物每日10:00—15:00补充UV-B 辐射,UV-B 灯为40 W南京牌UV-B灯管,最大峰值发射波长为320 nm(λ<320 nm),置于植株上方30 cm处。第2组每天辐射20 min(3.53 kJ·m?2·d-1),第3组每天辐射40 min(7.16 kJ·m?2·d-1),第4组每天辐射60 min(10.59 kJ·m?2·d-1),第5组每天辐射80 min(14.12 kJ·m?2·d-1)。用北京师范大学光电仪器厂生产的UV-B型紫外线辐射计测定UV-B强度。30 d后,成熟的番茄果实用于理化和感官分析。
1.2 果实形态和可溶性固形物含量测定
先用精密天平测量果实鲜质量(FM),然后在70 ℃的条件下干燥6 d,测量果实干质量(DM)。果实纵径和横径用电子数字卡尺测量,以mm表示。挑选红色成熟果实测定可溶性固形物含量。统计株高、结实节间长度、结实节数和每节果实数。
1.3 果实色泽和硬度的测定
用色度计(Minolta CR-300,美国)监测果实色泽。在番茄果实的中间区域进行测量,应用CIELaB色度系统分析果实颜色,结果用亮度、色度和色调表示。亮度在0(黑色)和100(白色)之间,色度值表示颜色饱和度,色调以度数表示,其中接近0°的是红色调,90°左右是黄色调,接近180°是绿色,270°是蓝色。应用德国STEPS水果硬度计测量番茄果实硬度,取3次平均值。
1.4 果实生物活性物质含量测定
抗氧化能力的测定:冷冻番茄果实干块(1 g)在30 mL V甲醇∶V去离子水=1∶1的溶液中提取,在室温下搅拌孵育30 min,搅拌均匀。提取液在10 000 r·min-1下离心10 min,收集上清液(SN),用提取液调节体积至30 mL,重复2次。最后,将上清液的体积调整为150 mL,用ABTS法测量抗氧化能力。
总酚含量的测定:采用福林酚法测定。(1)绘制标准样的处理和标准曲线。(2)番茄果实干物质中总酚含量的测定。取上述番茄果实干物质提取液1 mL于15 mL试管中,分别加入1 mL福林酚(FC)显色剂及3 mL 20% Na2CO3溶液,混匀,于50 ℃水浴反应30 min,用分光光度计在765 nm波长下测定吸光度,3次重复,将所测吸光度代入回归方程,计算总酚含量。
邻二酚类物质含量测定:采用钼酸盐法测定番茄果实中的邻二酚类物质,将5%(w,下同)钼酸钠溶液1 mL加入到4 mL的上清液中。在20 ℃下混合反应15 min,在370 nm处读取样品的吸光度,并计算邻二酚类物质含量。
总黄酮含量测定:按SILVA-BELTRA等[13]的方法测定总黄酮含量,略作修改。试验中用上清液250 μL、去离子水1.25 mL、亚硝酸钠溶液75 μL(5%)混合,5 min后加入150 μL的氯化铝溶液(10%),培养6 min后加入500 μL的氢氧化钠溶液 (1 mol·L-1)和250 μL的去离子水。用分光光度计在510 nm处读取吸光度,采用儿茶素标准曲线,结果用儿茶素的等价物表示。
1.5 参数统计分析
数据的方差分析使用SPSS 17.0(SPSS Inc,USA),采用單因素方差分析(ANOVA)对处理组与对照组进行比较。当数据有统计学差异时,采用Dunnett比较检验。
2 结果与分析
2.1 UV-B辐射对番茄果实数量、品质和外观的影响
结果表明,增加UV-B辐射对果实成熟同步性、果实数量和大小有显著影响,果实成熟同步性强,单株结实数量增多,果实体积减小。每天UV-B辐射20~40 min,即UV-B辐射剂量为3.53~7.16 kJ·m?2·d-1,30 d后,结实数量显著增多,由对照平均每株15.70个果实增加到平均每株20.30~21.80个果实,结实数量增加29.30%~38.85%。但随着UV-B辐射强度的继续加强,结实数量不再增加,反而与对照株结实数量相近(表1)。试验结果表明,增加UV-B辐射显著降低番茄单果质量,经过30 d UV-B辐射,剂量为3.53~7.16 kJ·m?2·d-1时,番茄单果鲜质量减少6.67%~8.33%,剂量为10.59~14.12 kJ·m?2·d-1时,番茄单果鲜质量减少33.33%。与单果鲜质量减少相似,经过30 d UV-B辐射,单果干质量下降,UV-B辐射剂量为3.53~7.16 kJ·m?2·d-1时,单果干质量下降5%~6.82%,剂量为10.59~14.12 kJ·m?2·d-1时,单果干质量下降27.27%~29.55%。在本试验条件下,UV-B辐射对番茄果实可溶性固形物含量影响最小,只有当辐射剂量大于14.12 kJ·m?2·d-1时才会导致番茄果实可溶性固形物含量显著下降(表1)。
由表2可知,增加UV-B辐射可显著降低番茄株高和结实节间长,且随着UV-B辐射剂量的增加,株高也随之降低。当UV-B辐射剂量增加7.16 kJ·m?2·d-1时,株高降低了14.1%。但其结实节数随着UV-B辐射剂量的增加而增加。低剂量UV-B辐射(3.53~7.16 kJ·m?2·d-1)并不影响每节果实数目,而番茄在高剂量UV-B辐射(10.59~14.12 kJ·m?2·d-1)下其每节果实数目显著减少。
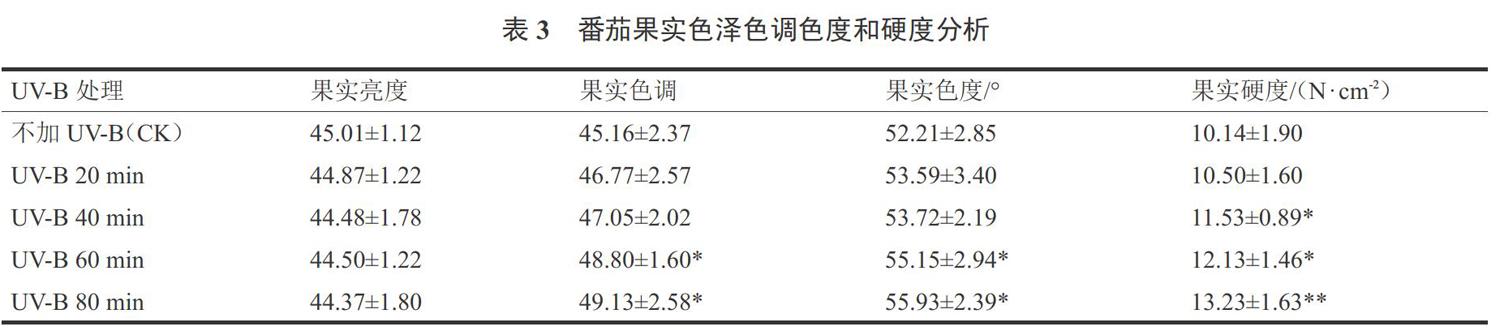
2.2 UV-B辐射对番茄果实色泽和硬度的影响
试验结果表明,UV-B辐射对番茄果实亮度没有显著影响。低剂量(3.53~7.16 kJ·m?2·d-1)的UV-B辐射对番茄色调和色度没有显著影响,高剂量(10.59~14.12 kJ·m?2·d-1)的UV-B辐射增加番茄色调和色度。温室种植番茄时,增加UV-B辐射有利于提高番茄果实硬度,当UV-B辐射强度达到7.16 kJ·m?2·d-1时,能显著增加番茄果实硬度,且果实硬度随着UV-B辐射强度的增强而增加(表3)。
2.3 UV-B辐射对番茄果实多酚类物质含量和抗氧化活性的影响
紫外线辐射损伤的表现形式之一是生物膜的过氧化损伤,类黄酮和类胡萝卜素等多酚类次生代谢物质是抗氧化剂,紫外线辐射能诱导植物产生多酚类次生代谢物质,从而增强植物抗氧化能力,减轻或防御紫外线对植物的损伤。
在温室番茄果实发育期增加UV-B辐射,30 d后收获成熟的番茄果实,测量其抗氧化活性及总酚、邻二酚类物质和总黄酮含量。由图1可知,低剂量(3.53~7.16 kJ·m?2·d-1)的UV-B辐射能提高温室番茄果实的抗自由基活性,而高剂量(10.59~14.12 kJ·m?2·d-1)UV-B辐射导致其抗自由基活性降低(a)。低剂量UV-B辐射使总酚含量增加,高剂量(10.59~14.12 kJ·m?2·d-1)UV-B辐射对总体含量影响不显著,还稍有降低(b)。低剂量(7.16 kJ·m?2·d-1)的UV-B辐射显著提高邻二酚类物质含量,而高剂量(14.12 kJ·m?2·d-1)UV-B辐射降低邻二酚类物质含量(c)。低剂量(3.53~7.16 kJ·m?2·d-1)的UV-B辐射显著提高总黄酮含量,高剂量UV-B辐射对总黄酮含量没有显著影响。
3 讨论与结论
太阳光是地球上植物的直接能量来源,是植物生长发育的重要环境因素,但是,太阳光谱中还有约5%的紫外线光波,根据波长不同分为紫外线A(UV-A,λ=320~400 nm)、UV-B(UV-B,λ=290~320 nm)、紫外线C(UV-C,λ=100~290 nm)。其中,UV-B生物活性最强,过强的紫外线辐射会使植物受到辐射损伤[14]。所幸大气的臭氧层对紫外线有很强的吸收作用,能吸收掉太阳光中的大部分紫外线,因此能保护地球上的生物,使它们免受紫外线的伤害。然而,随着工业发展,臭氧层有所减薄,从而导致地球表面紫外线辐射增强,因此,近几十年来,人们越来越担心太阳紫外线UV-B辐射可能会对植物生长产生负面影响[15]。然而,在自然生态系统中,由于多种环境因子间存在综合协同作用,UV-B辐射所造成的微妙的间接影响有时可能会比其对光合作用的直接影响更为重要[16]。在发现紫外线照射对植物细胞的作用之后,科学界也将紫外线视为人为调节植物生长发育、开花和结果的手段[17]。因此,在农业生产中,一种新的植物生长发育调控模式正在出现,这就是适度的紫外线补充可能改善作物营养价值(如改变植物次生代谢途径,改变次生代谢物的种类和含量),从而提高农产品作为健康食品的价值[18-19]。
本研究结果表明,增加UV-B辐射对温室番茄节间长度和株高、果实硬度、果实数量、果实大小有显著影响。从第一花序出现开始增加UV-B辐射,辐射强度为3.53 kJ·m?2·d-1,30 d后,节间长度缩短22.42%,辐射强度为7.16 kJ·m?2·d-1,节间长度缩短30.30%,由此可见,UV-B辐射能显著缩短节间长度,降低植株高度,且节间长度和株高与紫外线辐射强度成负相关。由于节间缩短,株高降低,在温室有限高度内,植株节间数增多,坐果节数增加,单株果实数量增加。但是,当紫外线辐射强度过大时(大于10.59 kJ·m?2·d-1),虽然节间长度进一步缩短,但单株结实数并不增加,这可能是強紫外线对植株造成伤害,光合效率下降,进而导致每节果实数量减少的原因(从正常平均每节4个减少到2个)。在番茄果实形成生长发育期每天UV-B辐射3.53~7.16 kJ·m?2·d-1,结实数量显著增多,果实体积略有减小。低强度紫外线照射组的果实略小、鲜质量和干质量略有降低,一方面是UV-B对植物光合作用有抑制现象,更直接的原因可能是结果数量增加。因此,生产中可采用大果品种以改善紫外线辐射导致果实体积减小现象,以便不影响其商品品质。但随着UV-B辐射强度的继续加强(10.59~14.12 kJ·m?2·d-1),结果数量不再增加,反而导致果实体积、单果鲜质量和干质量显著下降,显著降低了果实的商品品质。证明高强度的紫外线辐射不利于温室番茄生长和果实发育,因此,生产中只能用低强度的紫外线辐射。试验结果对其他温室作物种植也有一定指导意义,即通过适度增加UV-B辐射,增强果实成熟同步性和果实硬度,有利于果实收获和贮运,调控温室植株节间长度和植株高度,增加植株节数,从而增加果实数量。但是,因为每种植物对紫外线耐受性不同,确定具体植物适度的紫外线辐射强度尤其重要。
食用含有抗氧化能力的植物次生代谢活性物质的水果是消费者偏好的一个趋势,主要是因为它们在均衡饮食的同时具有多种健康益处[20-21]。UV-B照射诱导的次生代谢途径变化与抗氧化能力的活性物质产生积累密切相关[22]。但到目前为止,国内外有关UV-B照射的植物效应研究基本上着眼于防御紫外线的伤害,仅有少数报道涉及对果蔬品质的正向影响[23]。紫外线照射会改变植物的代谢物,即增加挥发性有机化合物的产生[24]。有研究报道,UV-B辐射刺激了紫苏的抗氧化能力[25]。在莴苣中,除去太阳UV-B后,花青素、黄酮类化合物和酚类物质比紫外线照射的植物减少,也反向证明紫外线辐射能诱导植物产生花青素、黄酮类化合物和酚类物质[26]。本试验结果表明,UV-B辐射使番茄果实酚类、邻二酚和黄酮类等抗氧化化合物含量显著升高,具有较高水平的抗氧化能力。本研究结果表明,补充UV-B对促进水果的抗氧化性能和增加某些次级代谢物的丰富度有很大好处。
总之,温室中适度增加UV-B辐射,可通过调控植株节间长度而增加番茄单株果实数量和果实硬度,诱导植物产生酚类、邻二酚和黄酮类等抗氧化化合物,增加果实抗氧化能力。温室果蔬生产中适度增加UV-B辐射是一种调控设施果蔬生长发育和改善品质的简便有效的技术方法。
参考文献
[1] 王欢欢,马越,白冰,等.番茄果实呈香组分及其代谢途径研究进展[J].中国瓜菜,2018,31(12):1-4.
[2] 刘怡玲,陈军,郑青松,等.5个樱桃番茄品种弱光下生长、产量及品质的比较[J].江苏农业科学,2017,45(21):158-161.
[3] 杨俊伟,梁婷婷,严露露,等.不同光质对番茄果实品质及挥发性物质的影响[J].食品科学,2019,40(11):55-61.
[4] 蒲晓宏,岳修乐,安黎哲.植物对UV-B辐射的响应与调控机制[J].中国科学:生命科学,2017,47(8):818-828.
[5] 荣瑞芬,冯双庆,赵玉梅.短波紫外线照射对采后番茄果皮中防御酶活性的影响[J].北京农学院学报,2002,17(4):89-92.
[6] WU D,HU Q,YAN Z,et al.Structural basis of ultraviolet-B perception by UVR8[J].Nature,2012,484:214-219.
[7] JENKINS G I.The UV-B photoreceptor UVR8:from structure to physiology[J].Plant Cell,2014,26:21-37.
[8] HAYES S,SHARMA A,FRASERD P,et al.UV-B perceived by the UVR8 photoreceptor Inhibits plant thermomorphogenesis [J].Current Biology,2017,27(1):120-127.
[9] CHRISTIE J M,ARVAI A S,BAXTER K J,et al.Plant UVR8 photoreceptor senses UV-B by tryptophan-mediated disruption of cross-dimer salt bridges[J].Science,2012,335(6075):1492-1496.
[10] 榮瑞芬,冯双庆.不同剂量短波紫外线照射对采后番茄后熟和发病的影响[J].中国农业大学学报,2001,6(1):68-73.
[11] WANG H B,GUI M Y,TIAN X,et al. Effects of UV-B on vitamin C,phenolics,flavonoids and their related enzyme activities in mung bean sprouts(Vigna radiata)[J]. International Journal of Food Science and Technology,2017,52(3):827-833.
[12] 董飞,王传增,孙秀东,等.基于蛋白质组学研究光质对番茄果实挥发性物质的影响机理[J].园艺学报,2019,46(2):280-294.
[13] SILVA-BELTRAN P,RUIZ-CRUZ S,CIRA-CH?VEZ L A,et al.Total phenolic,flavonoid,tomatine,and tomatidine contents and antioxidant and antimicrobial activities of extracts of tomato plant[J]. Scientia Horticulturae,2015,155:89-96.
[14] 郭建军,贵梦园,张佩佩,等.景天三七对紫外线B辐射敏感性研究[J].河南农业大学学报,2017,51(1):71-75.
[15] GUAN W,GAO K.Impacts of UV radiation on photosynthesis and growth of the coccolithophore Emiliania huxleyi(Haptophyceae)[J]. Environmental and Experimental Botany,2010,67(3):502-508.
[16] BERLI F J,MORENO D,PICOLI P,et al. Abscisic acid is involved in the response of grape(Vitis vinifera L. cv. Malbec)leaf tissues to ultraviolet-B radiation by enhancing ultraviolet-absorbing compounds,antioxidant enzymes and membrane sterols [J].Plant,Cell and Environment,2010,33(1):1-10.
[17] GUI M Y,HE H,LI Y D,et al.Effect of UV-B treatment during the growth process on the postharvest quality of mung bean sprouts(Vigna radiata)[J]. International Journal of Food Science and Technology,2018,53(9):2166-2172.
[18] CELINA D A,INACIO M D F,JONATHAN G.Instrumented color determination and sensory analysis of tomato fruits (Lycopersicum esculentum Mill.)[J].International Journal of Agricultural and Environmental Information Systems,2019,8(1):49-62.
[19] 王静,冯娜娜,王丹,等.UV-B 辐射对紫花苜蓿幼苗品质和抗氧化性的影响[J].食品科技,2016,41(6):56-60.
[20] MISHRA V,SRIVASTAVA G,PRASAD S M.Antioxidant response of bitter gourd (Momordica charantia L.) seedlings to interactive effect of dimethoate and UV-B irradiation[J].Scientia Horticulturae,2009,120(3):373 -378.
[21] CHICARO P,PINTO E,COLEPICOLO P,et al.Flavonoids from Lychnophora passerine (Asteraceae):potential antioxidants and UV protectants[J].Biochemical Systematics and Ecology,2004,32:239-243.
[22] XU C P,SULLIVAN J H,GARRETT W M,et al. Impact of solar Ultraviolet-B on the proteome in soybean lines differing in flavonoid contents[J].Phytochemistry,2008,69(1):38-48.
[23] 刘佳钰,王蓉,张弛,等.UV-B辐射与植物次级代谢产物变化的研究进展[J].基因组学与应用生物学,2017,36(8):3157-3166.
[24] 荣瑞芬,于涛.短波紫外线辐照对草莓贮藏保鲜效果的影响[J].食品科学,2003,24(1):146-149.
[25] 徐亚民,马越,赵晓燕,等.紫苏等4种天然色素抗氧化能力的比较[J].华北农学报,2007,22(2):187-190.
[26] 陈慧泽,韩榕.植物响应UV-B辐射的研究进展[J].植物学报,2015,50(6):790-801.
