转iaaM基因黄花蒿中基因表达量与其腺毛密度相关性分析
2020-04-04钟烨冰张学文
钟烨冰,肖 楠,胡 晴,黄 妤,赵 燕,张学文
(湖南农业大学生物科学技术学院,长沙 410128)
黄花蒿(Artemisia annua)是菊科蒿属的草本植物,植株含有挥发油、黄酮类化合物、香豆素、倍半萜类等物质[1]。其中倍半萜类的青蒿素类物质是一种新型特效治疗疟疾的药物,黄花蒿是青蒿素药物的主要来源[2]。黄花蒿作为抗疟药青蒿素的原料,在湖南、贵州、重庆、云南等省大面积种植,青蒿素类物质主要在其腺毛中合成和贮存,因此腺毛直接关系到黄花蒿中有效成分青蒿素的含量。近年来,经过对黄花蒿的品种改良,黄花蒿中青蒿素的含量得以不断提高。野生黄花蒿青蒿素含量在1~2 mg/g,当前栽培黄花蒿品种如新南方、A品系等的青蒿素含量已提高至10~20 mg/g,提高了10倍左右。但进一步提高黄花蒿中青蒿素含量,常规育种难有突破。采用现代生物技术对黄花蒿进行改良,则有望突破常规育种的局限。
来源于土壤根癌农杆菌的iaaM基因在植物中表达吲哚乙酸甲基化酶,能通过色氨酸途径合成生长素。利用iaaM基因开展植物遗传转化,可以调控其生长素合成并产生生长素相关的性状变异。尤其将该基因与植物特异启动子结合后在植物的特定组织器官表达,可研究生长素对植物器官形态发育的影响或改良作物农艺性状。王士杰等[3]将iaaM基因与棉花种皮特异表达基因在FBP7启动子调控下转化到棉花品系,略提高了纤维长度和比强度,且显著改良其纤维的马克隆值(p=0.0449*)而对其他性状没有产生不良影响。Zhao等[4]、彭彦等[5]将iaaM在拟南芥表皮毛特异表达Gl2基因启动子调控下转化烟草,能显著提高烟草表皮毛密度。龙炎杏[6]将iaaM基因在黄花蒿腺毛特异表达基因GTpro启动子调控下转化黄花蒿,也显著提高了黄花蒿腺毛的密度。说明特异表达iaaM基因能促进黄花蒿、烟草等植物的表皮毛和腺毛发育。但iaaM基因转化黄花蒿后基因的表达量与其腺毛细胞密度的相关性还缺乏系统性研究。笔者利用13个黄花蒿A品系独立转基因单株,开展了转基因定量表达与其腺毛密度及生长状况的分析。
1 材料与方法
1.1 材料
黄花蒿A品系由朱卫平提供,转iaaM基因的植株为龙炎杏等开展的根癌农杆菌叶盘共培法转化的再生苗,共13株,都为同一批次独立的转基因个体。转基因植株的基因表达水平会因叶片成熟度不同而有所差异,为减少这种误差,取材时,取每株材料从顶端分生组织起往下数不同成熟度的第2、12、22片叶片3片,鲜重50~100 mg。将以上3片叶片混合作为1个样本组提取其总RNA。
1.2 实验方法
1.2.1 基因表达的半定量
利用TransGen Biotech的试剂盒方法分别提取转基因单株、对照黄花蒿植株的总RNA。经紫外分光光谱检测和琼脂糖凝胶电泳检测其质量。
RNA反转录用Takara公司的PrimerScriptTMⅡ1st Strand cDNA Synthesis Kit试剂盒合成cDNA的第一链。反应体系为20μL:RNA 16μL和All MIX 4μL。将体系轻轻混匀,42℃孵育30 min,85℃加热5 s后终止反应。
基因表达定量以Actin作为内参基因,上游引物为:5′-ATGACATGGAGAAGATCTGG-3′、下游引物为:5′-CGCTCGGTAAGGATCTTCATC-3′,并根据iaaM基因的核苷酸序列设计特异性引物,上游引物为:5′-TCAGCGATAGAGAGGATCTTTCTGG-3′、下游引物为:5′-CGCCAATAGCTTGTGGGAGTC-3′,以cDNA为模板用特异引物进行PCR扩增,引物和金牌mix酶皆由湖南擎科生物技术有限公司合成和提供。内参进行半定量RT-PCR总体系25μL:上游引物0.5μL,下游引物0.5μL,金牌mix酶23.5μL,模板cDNA 0.5μL。调整转基因单株与对照的模板cDNA量,使内参基因Actin琼脂糖凝胶电泳的亮度一致,然后以该模板量扩增iaaM 基因。Actin反应程序条件:95℃预变性5 min,然后95℃变性30 s,58℃退火30 s,72℃延伸20 s为一个循环,循环25次,72℃最后延伸5 min。扩增目的基因的条件与上述的不同之处仅有退火为59℃,延伸45 s。
1.2.2 转基因黄花蒿腺毛数量统计
13个转基因单株和对照黄花蒿在营养土钵种植,生长至30 cm高时开始取样,单株共取不同发育时期的代表性叶片6片,每株材料从顶端往下数的第2、6、10、14、18、22片叶片,在奥林巴斯荧光显微镜下荧光观察并拍照,拍照软件为Motic Images Plus 2.0和Scion VisiCapture。在目镜10×、物镜4×下观察腺毛细胞,并拍摄成数码荧光显微照片。在数码照片上以6个0.09 mm2为单位的正方形随机排布,统计出不同转基因黄花蒿单位面积腺毛数量,并得出其平均密度。
1.2.3 数据分析
用Excel 2010进行统计,GraphPad Prism 7进行数据分析。
2 实验结果与分析
2.1 iaaM 基因的表达定量
以总RNA反转录后cDNA作为模板,扩增内参基因Actin,并根据Actin基因扩增情况进行模板量调整,对iaaM基因表达进行半定量分析,得到图1的结果。
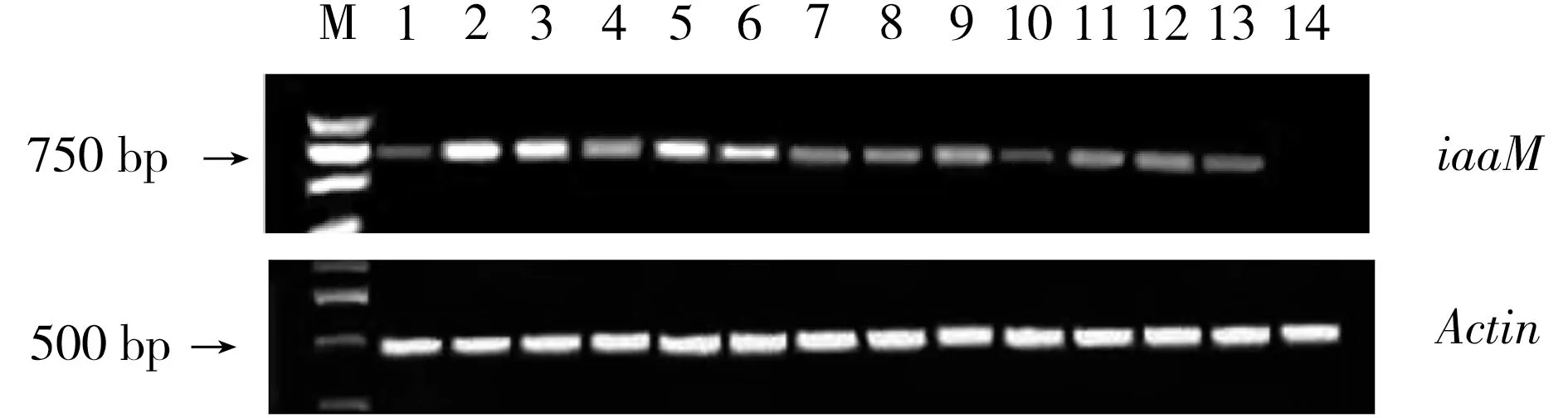
图1 iaaM 基因的半定量RT-PCR产物凝胶电泳图Fig.1 The agarose gel of semi-quantitative RT-PCR of iaaM
由图2可以看出,13个转基因单株中以iaaM-2、iaaM-3和iaaM-5基因表达量最高,iaaM-6和iaaM-4次之,iaaM-9、iaaM-7、iaaM-8、iaaM-11、iaaM-12号表达量接近,都比较低,iaaM-1、iaaM-10号和iaaM-13号表达量最低。
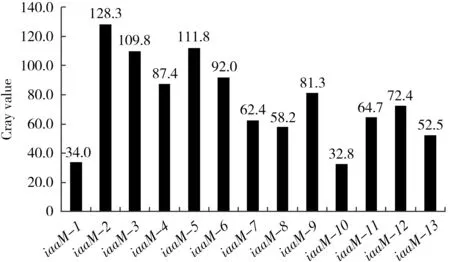
图2 iaaM 基因的半定量RT-PCR产物凝胶电泳灰度分析Fig.2 The Gray-scale analysis of semi-quantitative RT-PCR of iaaM by agarose gel
2.2 不同转基因个体腺毛细胞的密度观察
在植株同一生长期,每株材料从顶端分生组织起往下数取第2、6、10、14、18、22片叶片,共6片,在荧光显微镜下观察转基因黄花蒿与野生型黄花蒿腺毛细胞的分布情况,如图3。

按照设计的统计方法,选取6个同部位的单位面积,统计转基因个体的腺毛密度,结果如表1。

表1 转iaaM 基因黄花蒿与野生型个体腺毛密度统计Table 1 Statistical of glandular density in transgenic individuals compare with the control plantlet 枚/mm2
根据表1的统计结果,iaaM-2腺毛密度最高,相对野生型对照构成极显著差异;iaaM-3~iaaM-9、iaaM-11、iaaM-12对比野生型也达到极显著差异;iaaM-13对比野生型存在显著差异;iaaM-1和iaaM-10则无显著差异。对照转基因的表达定量分析结果,转基因植株腺毛密度与基因的表达量显然存在一致性;转基因黄花蒿腺毛密度都高于野生型黄花蒿;基因表达量越高的个体,腺毛密度也越高。野生型腺毛的平均密度为25.6枚/mm2,而转基因材料最高密度可达59.9枚/mm2。
3 讨论
外源基因在转基因植物中的表达量,直接影响着转基因性状的表现,也决定了其生产应用价值。在转基因实践中,基因重组构建的设计,包括启动子的强弱、阅读框架与密码子选择、顺式作用元件、翻译过程、表达系统等都影响外源基因在受体细胞中的表达[7~11]。本实验所使用的材料为青蒿腺毛特异表达GTpro启动子,与iaaM重组后转化黄花蒿,使基因在腺毛中表达。基因重组时整合位点不同是转基因个体间差异表达的重要原因之一。
根据谢德雄等在烟草中的研究,iaaM基因在表皮毛特异启动子调控下的转化,促进了烟草表皮毛发育,是由于叶片表皮细胞的早期分化调控存在的反馈调节[12]。生长素促使叶片表面的细胞有更多向表皮毛发育的倾向,因而提高了表皮毛的密度。黄花蒿的转化也验证了利用iaaM基因的效果。利用腺毛特异表达启动子调控生长素合成,也能促进黄花蒿腺毛的发育,提高腺毛的密度。
本实验采用半定量的方法对基因表达量进行分析,这种方法对其定量有一定的局限性,但也能很直观地通过凝胶中分子荧光亮度判断出相对量大小。腺毛密度的观察,转基因黄花蒿腺毛密度与基因表达量成明显的正相关性,说明腺毛细胞的发育启动也存在着生长素与该特异启动子的反馈作用。不同的转基因个体因为转基因整合位点的不同,使GTpro调控的iaaM有了差异化的表达水平。
启动子特异性是决定iaaM基因表达对植物发育影响的关键。彭彦[13]利用韧皮部特异表达启动子在拟南芥中过量表达iaaM,观察到植株出现下胚轴及叶柄变细增长、成熟植株相对矮小、叶片窄小向下卷曲等现象。在转基因烟草中同样出现这种异常表型,如茎上出现大量不定根、植株矮小、叶窄小卷曲且厚度增加等。文红秀[14]利用组成型表达启动子在油菜中过表达iaaM,出现了植株较野生型矮、每株角果数、每果粒数、千粒重等增加的现象。
因此,要利用生长素合成基因来改良植物性状,关键在于对该基因的表达调控,用特定的启动子在特定的组织细胞中提高生长素的含量,才能有效改进其农艺性状。本实验通过转iaaM基因表达量与黄花蒿腺毛密度的相关性分析,证明转基因在植株个体中表达量各有差异,而且其表达量与腺毛密度存在着正向关联,可为选育高青蒿素含量的黄花蒿新品系提供参考。
