福建柏优良单株自由授粉子代测定及选择
2020-03-31苏顺德
苏顺德
(福建省林业科学研究院、国家林业局南方山地用材林培育重点实验室、福建省森林培育与林产品加工利用重点实验室,福建 福州 350012)
福建柏(Fokieniahodginsii(Dunn) Henry et Thomas)为柏科(Cupressaceae)福建柏属唯一种,被《中国物种红色名录(第一卷):红色名录》列为易危(VU)物种[1],主要分布在中亚热带湿润气候区,介于22°—28°30′N、102°—120°E间[2-3],是分布区内常绿阔叶和针阔混交天然林的重要组成树种[4]。福建柏具有生长快、耐瘠薄、树干通直,木材纹理直、结构细、切面光滑、胶粘性良好的特点,是建筑、装饰装璜、雕刻的优良用材[2]。由于过度砍伐,福建柏天然资源极度匮乏[1],但在中国南方林区广泛栽培[5]。中国于1996年将福建柏育种研究列入国家“九五”林业科技攻关项目,重点开展的全分布区种源试验发现福建柏生长在种源水平受到高度的遗传控制,幼林期树高、胸径的广义遗传力达到0.6以上[6-8]。在现存天然育种资源遗传多样性不高的背景下[3-4],如何利用人工林种质资源,广泛构建福建柏选择群体,提高可利用种质的遗传多样性和目的性状的优良性,有力推进福建柏育种进程,支撑福建柏人工林建设成为福建柏遗传改良面临的重要问题。因此,福建省林业科学研究院在前期种源试验基础上,利用从1998年营建的福建柏种源试验林[9]中筛选的优良单株的自由授粉家系开展子代测定,估算这些优良单株生长性状的育种值和基因型值,据此筛选速生家系和无性繁殖原株,以充分发掘保存的福建柏种质资源的应用潜力。
1 材料与方法
1.1 试验点概况
试验点位于福建省邵武卫闽国有林场、福建省沙县官庄国有林场和福建省大田梅林国有林场,均属亚热带季风气候。各点试验地概况见表1。
1.2 试验材料与方法
根据1998年营建在福建省南靖国有林场和福建省永春碧卿国有林场的福建柏种源试验林、种源内家系试验林[7]的生长性状遗传变异分析结果[8],于2008年从中筛选速生种源、家系中的优良单株采集自由授粉种子,2009年春育苗,2010年春在福建省邵武卫闽国有林场、福建省沙县官庄国有林场和福建省大田梅林国有林场营建自由授粉子代测定林。参试家系均为相同的24个,以福建省安溪白濑国有林场(现福建省安溪丰田国有林场)初级种子园混合种为对照(表2)。田间试验为完全随机区组设计(RCB),25个处理,10次重复,4株单列小区。株行距2.0 m×2.0 m,种植穴规格50 cm×40 cm×30 cm。根据《福建柏丰产林培育技术规程》[10]管理试验林,但未修枝和间伐。试验林1~3 a林龄时每木调查树高,4~9 a林龄时每木调查树高、胸径,每次调查记录枯死株。

表2 参试家系及其母本种源
1.3 统计分析方法
根据树高、胸径估算单株立木材积[11]:V=0.00005685D1.629996H1.261954。根据枯死株数量计算每个小区保存率:保存率=[(4 -小区枯死株数量)/4]×100。
2 结果与分析
2.1 保存率及生长性状表型分析
由连年保存率曲线(图1)可知,造林当年试验林平均保存率为87.70%,第3年下降到84.85%,第4年降低到83.68%后逐年稳定在83.58%。由连年生长量曲线(图2)和平均生长性状表现(表3)可知,试验林平均树高连年生长量在2~6 a林龄时较为稳定,维持在0.82~0.89 m之间,7 a林龄时下滑至0.52 m,9 a林龄时又回升至0.94 m,且总生长量达到6.94 m。胸径连年生长量在6 a林龄时达到最大值1.68 cm,7 a林龄时开始下滑,9 a林龄时下滑至0.90 cm,且总生长量为9.79 cm。单株立木材积连年生长量在8 a林龄前稳步小量上升,9 a林龄时迅速上升至0.00724 m3-且总生长量达到0.02796 m3。树高、胸径和单株立木材积在单株间的分化虽然随着林龄增加而减小,但9 a林龄时表型变异系数仍分别高达11.80%、23.83%和45.11%,单株立木材积表型变异最为丰富,其次是胸径。

表3 试验林平均生长性状
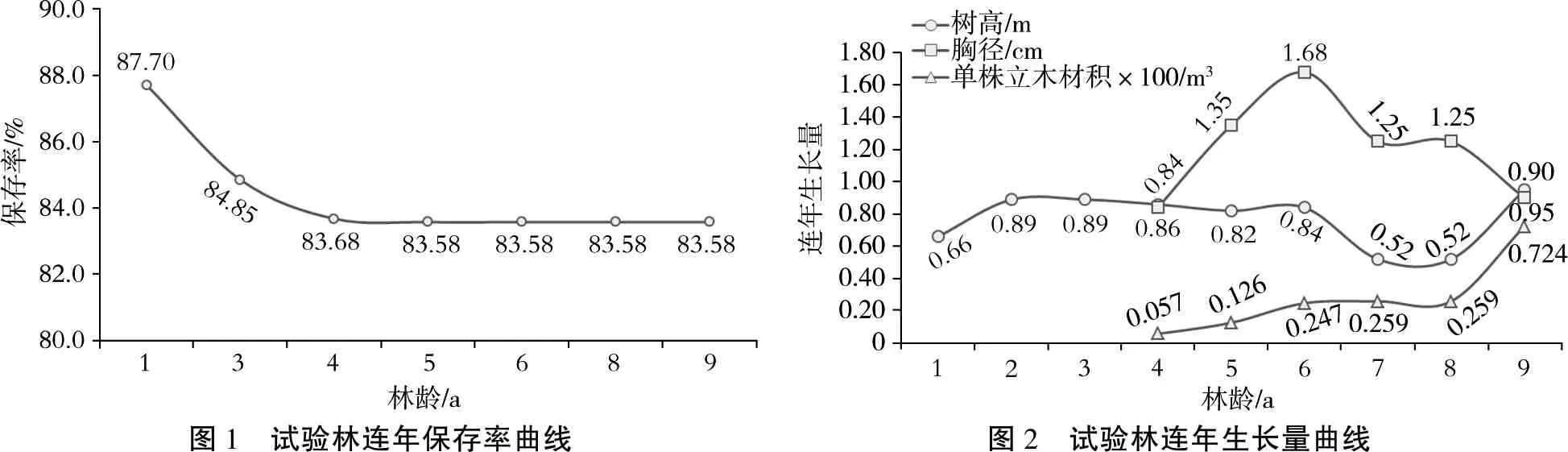
图1 试验林连年保存率曲线图2 试验林连年生长量曲线
2.2 保存率遗传变异分析
保存率在家系间差异极显著,试验地点与家系的交互作用也达显著水平(表4)。广义遗传力和狭义遗传力均在4 a林龄时出现最大值(图3),分别达到0.3675和0.0976,而此时正是保存率均值趋于稳定的林龄(图1)。之后广义遗传力较为稳定地介于0.2444~0.3105之间,狭义遗传力较为稳定地介于0.0573~0.0764之间。总体来看,广义遗传力明显高于狭义遗传力,表明保存率的加性遗传效应较小,而非加性遗传效应较为丰富。因此,在利用广义遗传力选择无性繁殖原株时,兼顾保存率可最大限度获得适应性方面的遗传增益;而在利用狭义遗传力选择家系时,可忽略家系间保存率的遗传差异,仅要求家系保存率具正向遗传增益即可。

表4 保存率方差分量及遗传力
*:**为差异极显著;*为差异显著;下同。
2.3 生长性状遗传变异分析
从表5—表7的生长性状方差分量估算结果可知,树高、胸径和单株立木材积在家系间差异均极显著,试验地点与家系的交互作用、试验地点与家系与区组的交互作用均达极显著水平。结合图4的遗传力随林龄变化趋势看,树高的广义遗传力在0.6864~0.7933间轻微波动,胸径的广义遗传力在0.7347~0.7688间轻微波动。单株立木材积的广义遗传力波动较大,从6 a林龄时迅速上升(从5 a林龄时的0.1476上升至6 a林龄时的0.5359),8 a林龄时开始稳定在0.7620~0.7641间。生长性状的狭义遗传力随林龄波动较小,特别是6 a林龄后树高狭义遗传力介于0.1362~0.1941间,胸径狭义遗传力介于0.1752~0.1756间,单株立木材积狭义遗传力介于0.1831~0.1887间。虽然5 a林龄前保存率遗传力波动较大,但树高、胸径的广义遗传力和狭义遗传力波动均较小,间接表明了生长性状和保存率间的遗传独立性。当利用8 a林龄前树高、胸径的广义遗传力进行早期筛选无性繁殖原株时,由于单株立木材积广义遗传力的不稳定性,存在较大的误选及漏选风险;而利用树高、胸径的狭义遗传力筛选优良家系却是可行的,但林龄宜在6 a以后。
2.4 B型遗传相关分析
试验点间各性状B型遗传相关系数—林龄曲线(图5)表明各林龄阶段,树高、胸径B型遗传相关系数均较高,达到0.881以上,6 a林龄以后均达到1.0,即6 a林龄后,参试家系树高、胸径在各试验点间的排名高度遗传相关,基因与环境互作较小。单株立木材积B型遗传相关系数呈明显的逐年升高趋势,林龄8 a后达到1.0,保存率B型遗传相关系数均较小,基因与环境的互作对保存率具有较大的影响。因此,在8 a林龄后选择能获得生长性状更为稳定的优良无性繁殖原株和家系,鉴于保存率广义遗传力相对较高,筛选无性繁殖原株时应该兼顾保存率的表现。

表5 树高方差分量及遗传力

表6 胸径方差分量及遗传力

表7 单株立木材积方差分量及遗传力
2.5 无性繁殖原株及家系选择
在以上遗传变异分析基础上,根据9 a林龄时基因型值估算的无性系遗传增益排序从参试的24株优良单株(1998年营建的种源试验林和种源内家系试验林中的单株)中选择无性繁殖原株,为进一步开展无性系测定、筛选优良无性系奠定基础。同时,利用9 a林龄时育种值估算的家系遗传增益排序从参试家系中筛选优良家系,充实福建柏选择群体,为构建育种群体、生产群体奠定基础。
以平均单株立木材积遗传增益不低于30%,保存率遗传增益不低于5.0%为指标筛选出无性繁殖原株11株(表8),树高、胸径、单株立木材积和保存率均值分别为7.09 m、10.18 cm、0.03133 m3和87.28%,遗传增益分别为6.84%、12.57%、30.07%和5.74%。以平均单株立木材积遗传增益不低于7%,保存率具有正向遗传增益,兼顾种源,筛选出优良家系8个(表9),树高、胸径、单株立木材积和保存率分别为7.10 m、10.22 cm、0.03162 m3、88.87%,遗传增益分别为1.47%、3.02%、7.80%和1.47%。

表8 无性繁殖优良原株9 a林龄生长状况

表9 优良家系9 a林龄生长状况
3 小结与讨论
福建柏人工纯林太阳能转化效率为1.42%,能量流动速率较高,与杉木人工纯林相比,能量年净增量较大,凋落物养分周转快,对地力维护极为有利[14-15]。14 a林龄的福建柏杉木混交林地上、地下空间分布成层性明显,养分空间得以扩大,而福建柏纯林地被物吸持水量达到32.09 t·hm-2,土壤中有机质、全氮、速效氮等含量均较杉木纯林高[5]。与中国南方广泛栽培的用材树种杉木相比,福建柏木材的气干密度、干裂势、硬度等表现较优[16]。已有研究表明福建柏生长速度快,30 a林龄人工林树高、胸径连年生长量仍然达到0.26 m、0.37 cm[17]。因此在森林生态功能维护和优质木材生产方面,福建柏在其产区均是优良的造林树种,发展潜力巨大,良种需求旺盛。但目前福建柏遗传改良水平相对较低,还处在第1代(轮)育种末期[8,18-19]和第2代(轮)育种起步阶段[20],良种供应不足。在遗传资源相对较为匮乏的现实情况下[3],如何充分利用已建立的种源试验林等基本群体,有效构建选择群体和育种群体,是福建柏高世代育种面临的关键问题。本研究在前期种源试验基础上,对种源试验林,种源内家系试验林中优良单株进行自由授粉子代测定,一方面筛选优良家系,另一方面选择无性繁殖原株,在充实福建柏选择群体的同时,为福建柏无性繁育奠定基础。试验结果表明,虽然保存率、树高、胸径和单株立木材积在家系间差异均极显著,但均受到较弱的遗传控制。9 a林龄前,保存率狭义遗传力维持在0.0389~0.0976间,树高狭义遗传力维持在0.1362~0.1941间,胸径狭义遗传力维持在0.1752~0.1756间,单株立木材积狭义遗传力维持在0.1831~0.1887间。福建柏优良单株家系间生长性状的遗传差异低于种源[8]但高于初级种子园自由授粉家系[18]。B型遗传相关分析表明在8 a林龄后选择能获得生长性状最为稳定的优良家系和无性繁殖原株。因而依据参试家系9 a林龄时遗传增益排序从参试的24株优良单株中选出11株作为无性繁殖原株,树高、胸径、单株立木材积和保存率均值分别为7.09 m、10.18 cm、0.03133 m3和87.28%,遗传增益分别为6.84%、12.57%、30.07%和5.74%。同时筛选出8个速生优良家系,树高、胸径、单株立木材积和保存率分别为7.10 m、10.22 cm、0.03162 m3和88.87%,家系遗传增益分别为1.47%、3.02%、7.80%和1.47%。
利用依据基因型值选择的无性繁殖原株经过无性化后开展无性系测定,并筛选优良无性系,无疑有效提高了福建柏无性系选择的效率和准确性,为解决短期内福建柏良种壮苗供需矛盾具有现实意义和重要作用。而依据育种值筛选的优良家系则是福建柏选择群体或者种质资源的重要组成部分,是进一步构建育种群体的关键材料。
