杉木半同胞子代胸径变异和大径材家系选择
2020-10-09邓厚银胡德活林军赖旭恩邓文剑韦如萍王润辉晏姝郑会全
邓厚银, 胡德活, 林军, 赖旭恩, 邓文剑, 韦如萍, 王润辉,晏姝, 郑会全*
(1. 广东省森林培育与保护利用重点实验室/广东省林业科学研究院,广州 510520;2. 乐昌市龙山林场,广东 乐昌 512221)
杉木(Cunninghamia lanceolata)是中国亚热带地区最主要的针叶用材树种[1]。自20世纪60年代以来, 该树种一直被视为长期育种的对象,旨在提高其速生性和适应性[2]。目前,杉木良种选育方面已取得较好的成效,而良种的使用,加上集约化的营林管理,杉木商品林轮伐期极大缩短并向短轮伐期优质林建设方向发展,但所产木材也以中小径材林为主,大径材产出少[3-4]。近年来,国内外市场对大径材的需求与日俱增,如何高效培育大径材已成为杉木商品林建设研究的一个热点。
通常认为,采用遗传控制、立地控制、密度控制、施肥管理等综合配套技术措施可实现大径材的定向培育[5-8]。有研究特别提出,杉木大径材定向培育良种效应尤为重要[6,8-9]。随着杉木高世代良种选育进程的推进,“十三五”期间国家科技部还启动了国家重点研发计划“杉木高效培育技术研究”项目课题“杉木大径材高效培育技术研究”,针对高世代育种背景下的杉木大径材培育系列问题展开科技攻关。有鉴于此,本研究以广东杉木第二代种子园半同胞子代(28个半同胞家系)为对象,对其多年度的胸径情况(林龄为4、9、11、16和21 a生)进行持续测定分析,探讨其胸径变异规律并筛选出具有大径材培育潜力的候选家系,进而为杉木大径材优良家系选育研究提供参考。
1 材料和方法
1.1 试验地概况
试验地位于广东省韶关乐昌市龙山林场(25°05′N,113°27′ E),属中亚热带气候区,年降雨量1 550~1 750 mm,年均温19.6℃,土壤为赤红壤[10]。
1.2 材料
以杉木(Cunninghamia lanceolata)第二代种子园亲本子代为材料,包括28个半同胞家系(F3~ F30),并以第一代种子园(初级园,C1)和第一代改良种子园(1.5代园, C2)混合种子代为对照,于1998年2月9日营建子代测定林(造林苗木均为1 a生子代苗木)。测定林采用完全随机区组设计,16个区组,4株小区,株行距2 m×2 m。
1.3 方法
在林龄为4、9、11、16和21 a生时对杉木每木胸径进行测量。按照广东省地方标准DB44/T 1532-2015《杉木大径材培育技术规程》的大径材(树龄20 a以上,胸径≥25 cm的原木)标准,并参考相聪伟等[11-12]的方法设定立木径级:胸径≥25 cm为大径材、16~25 cm为中径材、6~16 cm为小径材(包括小条木)、<6 cm为薪材。
采用Excel 2010对数据进行整理与分析。方差分析(ANOVA,F值)、家系遗传方差分量(σg2)、家系遗传力(Hf)、单株遗传力(Hi)、年度遗传相关系数、家系育种值(BVf)计算采用南京林业大学童春发教授[13]开发的基于R系统的HalfsibMS和HalfsibBV软件进行,所用模型为单地点混合模型:yijk=μ+Bi+Fj+BFij+eijk,式中,yijk为第i个区组里第j个家系内第k个单株的数量性状观测值;μ为总体平均值;Bi为第i个区组的效应;Fj=为第j个家系的效应; BFij为第i个区组里第j个家系的交互效应;eijk为随机误差效应。表型变异系数估算采用公式CV(%)=100×S/,式中S和分别为测定值标准差、群体均值;遗传变异系数(GCV)估算采用公式GCV(%)=, 式中,σg2和分别为遗传方差分量、群体均值。21 a生时的材积[14]计算公式为V=0.000058777042×D1.9699831×H0.89646157,式中,D和H分别为胸径和树高。
2 结果和分析
2.1 胸径生长模式和家系间的差异
杉木第二代种子园半同胞家系(F3~F30)和对照家系(C1、C2)的胸径生长模式见图1,林龄4 a生时的平均胸径为6.3 cm,5 a后(至林龄9 a生)增幅达8.0 cm,至林龄11 a生时平均胸径为15.8 cm,5 a后(至林龄16 a生)胸径增幅明显放缓(仅为2.4 cm),而再过5 a后(至林龄21 a生)的增幅更低(仅1.9 cm)。就年均生长来看,林龄4和9 a生的年均胸径生长速率较高且相近(约1.6 cm/a),到林龄11、16和21 a生时,年均胸径生长速率偏低,且随树龄的增长有下降趋势(分别为1.4、1.1和1.0 cm/a)。
对比不同家系(包括对照家系)可见,不同家系相同林龄的胸径生长表现不一(图1)。各林龄家系间的胸径差异均达极显著水平(P<0.01),其中林龄4 a生的差异最大,其表型变异系数(CV)为6.29%,胸径生长最快家系是最慢家系的1.3倍(表1)。
2.2 胸径的遗传参数
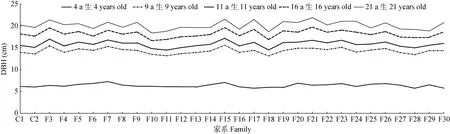
图1 不同林龄杉木家系的胸径(DBH)Fig. 1 Diameter at breast height (DBH) of Cunninghamia lanceolata families at different ages

表1 杉木家系的胸径(DBH)Table 1 Diameter at breast (DBH) of Cunninghamia lanceolata families
对杉木家系不同林龄的胸径遗传参数进行分析(表2),林龄4 a生时,家系的胸径遗传方差分量(σg2)和家系遗传力(Hf)分别为0.046 1和0.335 9,且随年龄的增长均呈增加趋势,至林龄21 a生时σg2达到0.529 9,Hf在林龄16 a生时达到0.548 6,随后略有下降(林龄21 a生时为0.544 2)。单株遗传力(Hi)同样随林龄的增大而增大,林龄4 a生时为0.059 3,至林龄16 a生时达到0.114 6,但林龄21 a生时有所降低。值得注意的是,相同林龄杉木家系的Hf均显著高于Hi,约高出4.78~5.66倍。另外,家系遗传变异系数(GCV)在不同林龄间差异较小(2.95%~3.62%),最小GCV出现在林龄9 a生时。
多年度遗传相关分析结果还表明(表3),林龄4 a生时杉木家系的胸径与其余林龄的相关关系呈随林龄增加而不断弱化的趋势,其中与林龄9 a生时的相关关系最为密切,遗传相关系数达0.631 9,而与林龄21 a生时的相关性降至0.115 8。林龄9、11、16和21 a生间的胸径遗传相关关系极高且较为稳定,相关系数均大于0.800 0。
2.3 家系胸径育种值的估算
以胸径为目标,对参试的28个半同胞家系和2个对照家系的不同林龄时育种值进行估算并排序,结果表明,各家系按育种值大小排名因林龄不同而有所变化(图2),但综合各年度胸径育种值(均值)可知,F3、F4、F5、F7、F9、F15、F17、F19、F20、F21、F22、F23、F24、F26、F30等15个家系排名靠前,且明显高于对照家系C1和C2。其中,有8个家系(F3、F5、F7、F9、F15、F21、F23和F26)各林龄的育种值排名均稳定在15名以内(50%入选率),尤其是这8个家系综合育种值排名还均在前11名内(具体排序为:F21>F15>F23>F3>F7>F26>F9>F5),显然具有更大育种潜力。

表2 杉木家系的胸径遗传参数Table 2 Genetic parameters of DBH among Cunninghamia lanceolata families

表3 杉木家系不同林龄间的胸径遗传相关系数Table 3 Genetic correlation coefficient of DBH of Cunninghamia lanceolata families among different ages
2.4 杉木家系各径级立木占比及大径材家系选择
鉴于参试家系林龄已达21 a生,适于立木径级类别划分。对各家系的大径材、中径材、小径材(包括小条木)、薪材立木(子代)占比进行分析(图3),结果表明不同家系的各径级立木占比不一致。大径材、中径材、小径材(包括小条木)、薪材立木占比分别为5.6%~29.2%、47.9%~73.5%、8.1%~34.0%和0~2.1%,其中以大径材立木占比的变化幅度最大。而家系F5、F9、F21大径材立木占比在参试家系中较高(排名前3),分别达到29.2%、27.5%和28.6%,且家系平均生长量也较为突出(前11名:胸径≥20.5 cm、材积≥0.310 9 m3)。值得注意的是,这3个家系也是“具有更大育种潜力”的家系,表明F5、F9、F21是参试家系中培育大径材的优良家系。
3 结论和讨论

图2 杉木家系随不同林龄期的胸径育种值排名Fig. 2 DBH breeding value rank of Cunninghamia lanceolata families at different ages

图3 21 a生杉木家系的径级立木占比Fig. 3 Ratio of volume at different diameter grade of Cunninghamia lanceolata families at 21 years old
胸径关乎木材产量,也是选育大径材良种的首要指标。本研究分析了广东杉木第二代种子园半同胞家系及其对照家系多年度的胸径生长模式, 结果表明,同一林龄的家系间胸径差异均达极显著水平,胸径表型变异系数(CV)为4.43%~6.29%,而胸径遗传变异系数(GCV)为2.95%~3.62%。胸径受较高遗传控制(Hf=0.335 9~0.548 6),且Hf随年龄的增长呈增加趋势,至林龄16 a生时达到0.548 6,随后趋缓。林龄9 a生时杉木胸径与早期(林龄4 a生)、晚期(林龄11、16、21 a生)胸径的遗传相关性最为密切。基于多年度育种值和林龄21 a生时家系的径级立木占比统计结果,共选择出F5、F9、F21等3个适于培育大径材的优良家系。
掌握林木胸径生长规律是开展良种选育和应用工作的基础。伍汉斌等[15]以中国199个杉木种源为材料,通过区域测定分析了33 a内5个林龄期参试材料的生长表现,认为不同种源杉木胸径在各林龄期的差异极显著,且具有明显的地理变异规律,胸径遗传力在生长前期(前12 a)呈逐年增加但后期渐稳趋势,同时胸径年度遗传相关较高,这为杉木优良种源胸径生长研究提供了参考。而戴俊等[16]以中国10个省区14个杉木无性系种子园84个半同胞家系为试材,获得了30 a间4个林龄段的胸径生长变异信息,认为家系胸径遗传力总体较高,早期(前5 a)胸径与后期的相关性随林龄增加呈不断弱化趋势,林龄8 a生的杉木胸径与前(林龄5 a生)后(林龄26和30 a生)期的相关性较为突出。本研究探讨了广东杉木第二代种子园半同胞家系的胸径生长变异规律,家系间的胸径差异极显著,胸径的家系遗传力较高且具有明显的年龄效应。值得注意的是, 单株遗传力与家系遗传力的变化趋势相近,这为参试家系的胸径生长选择及优良家系优良单株协同选择提供了依据。事实上,Zheng等[17]针对福建省杉木第3代育种群体半同胞家系的研究结果也表明,家系遗传力和单株遗传力在一定生长期内(林龄4~9 a生)均呈逐年增加趋势。但陈瑞生等[18]以杉木多系授粉子代为对象估算的家系遗传力和单株遗传力则没有明显的逐年增加趋势。此外,陈孝丑等[19]分析了杉木第2代种子园双列杂交子代的生长变异情况,估算的胸径家系遗传力和单株遗传力均随林龄增加呈下降的趋势。而在南亚热带乡土阔叶树种的研究上,朱积余等[20]对红锥(Castanopsis hystrix)家系的遗传变异研究表明,胸径的家系遗传力和单株遗传力均会随林龄的增大而逐渐减小,到7~9 a生时趋于稳定。这些差异可能与研究对象类型不同有关。
另外,本研究以较规律的测定年度(包括3个5年期:林龄4~9、11~16和16~21生)系统分析了杉木半同胞家系的胸径年度遗传相关性,结果表明,早期(林龄4 a生)的胸径与其余林龄期的相关关系呈随林龄增加而不断弱化趋势,与戴俊等[16]的研究结果相似。而毛红等[21]以16 a为尺度探讨了杉木家系年度胸径生长的相关性,也有类似结果。这说明早期生长差异或许不能够很好地反映出后期(尤其是16 a以后)的变化情况,家系间差异性可能会随林龄的变化而变化。事实上,育种值估算与排序结果也已证实,不同家系胸径大小排序会随林龄的变化而变化。值得关注的是,林龄9 a生时的杉木胸径与早期(林龄4 a生)、晚期(林龄11、16和21 a生)胸径的遗传相关性均尤为密切(P<0.01),遗传相关系数均大于0.80,这明显高于林龄4 a生的胸径与后期的遗传相关系数(0.631 9、0.475 5、0.216 9、0.115 8)。特别是,林龄9 a生与21 a生的胸径遗传相关系数远高于4 a生与21 a生时的遗传相关值(0.801 5 vs 0.115 8)。因此,林龄9 a生可能是开展后期大径材家系选择的重要时期。
曾志光等[9]认为,使用优良种源培育大径材比普通种源材积生长提高30%左右,而从优良种源中选育出的优良无性系培育的大径材,在优良种源的基础上又提高20%以上,且林分大径材比例明显增加。叶功富等[22]对12个产地65个杉木优良家系进行造林对比试验,认为选用优良家系可以缩短杉木大径材培育年限。这说明杉木大径材的定向培育亟需适宜的种源、家系或无性系。为选育适宜杉木大径材定向培育的种质,郑会全等[23]对杉木初级种子园23 a子代林大径材(胸径≥26 cm)优树进行选择,所选单株胸径现实增益均可超过56%,具有较大的应用潜力。而本研究在大径材适选年龄期(林龄21 a生)时对不同家系各径级立木占比尤其是大径材立木占比进行统计分析,并结合多年度育种值计算结果,共选择出3个适于培育大径材的优良家系,大径材立木占比均在27.5%以上,这些家系为杉木大径材的高效培育提供了候选种质来源。
