如何对濒危物种进行评估与拯救?*
2020-03-27谢平
谢 平
(1:中国科学院水生生物研究所东湖湖泊生态系统试验站,武汉 430072)(2:云南大学生态与环境学院高原湖泊生态与治理研究院,昆明 650500)
物种并不是永生的,地质历史上的绝大多数物种都已灭绝. 譬如,中国东部始新世时期的化石鱼类——鲤科、亚口鱼科、鲿科和鲈形目等都已绝灭[1]. 物种灭绝是一种自然过程,历史上曾经灭绝的许多物种皆是如此,但这是一种漫长的过程,通常是在数十万年甚至更长的时间尺度上发生. 物种自然灭绝的原因应该是多种多样的,但真正能够确认的并不多,譬如恐龙为何灭绝至今都没有定论.
如果一个物种正在走向灭绝之道,就称为濒危物种(endangered species). 现代物种的濒危绝大多数起因于人类活动的干扰,譬如,栖息地的改变、破坏或丧失、过度捕杀以及环境污染等. 根据世界自然保护联盟(International Union for Conservation of Nature,简称IUCN)2012年的报告,全球共有3079种动物和2655种植物处于濒危状态.
在自然界,一些物种缓慢的灭绝,更多的物种缓慢的诞生. 但是,人类的繁荣在很短的时间内给地球环境带来了巨大的变化,譬如,数以百万计的生物物种的栖息之地正在被人类挤压或占用,这些物种正在以惊人的速度灭绝或正在绝灭之中,但新物种却完全来不及产生.
非常遗憾的是,我们眼睁睁地看着很多物种灭绝或走上了不归之路,等我们意识到的时候,往往已回天乏术. 长江的白鱀豚和白鲟(功能性)灭绝了,中华鲟也无力回天,江豚也正在垂死挣扎之中……而我们甚至不知道这些悲剧的真正原因,一些词藻华丽的保护生物学理论经常是南辕北撤. 对物种生存状态(特别是濒危状态)的诊断与评估是物种保护与拯救的关键一步.
1 濒危物种评估发展简史
人类对物种濒危的关注可追溯至19世纪中叶的保护运动(conservation movement),由于农业的扩张和狩猎工具的进步,一些对英国文化和生态均具有重要意义的动物,如大鸨(Otistarda)、海雕(Haliaeetusalbicilla)和大海雀(Pinguinusimpennis)等相继灭绝,这催生了世界上第一部自然保护法——1869年英国海鸟保护法案诞生[2].
进入20世纪,物种面临的形势日趋严峻. IUCN于1948年正式成立,并于1963年开始编制濒危物种红色名录(Red List of Threatened Species). 1992年,在巴西里约热内卢举行的联合国环发大会上签署的《生物多样性公约》明确了各国政府对物种保护的法律义务[3].
濒危评估的第一步就是对濒危状态进行等级划分,这种等级能够反映物种绝灭的发生机率,如极危>濒危>易危,不难理解,极危意味着物种灭绝的几率极高.
在IUCN 2012年公布的名录中,一共有3079种动物和2655种植物处于濒危状态. 许多国家制定了法律来保护濒危物种,如禁止捕猎、限制土地开发或建立保护区等.
生物多样性丧失与保护,特别关注受威胁程度评估与物种红色名录和生态系统红色名录、物种灭绝速率、保护空缺分析、现状与保护进展评估指标等依然是现在生物多样性研究的热点[4].
但是,如何通过一定的指标对物种的濒危程度进行定量评价却并不那么容易. 即便如此,人们还是提出了一些判定标准.
2 物种濒危标准体系
1991年,英国学者梅斯(G. Mace)和兰德(R. Lande)提出了根据在一定时间内物种的灭绝概率来确定物种濒危等级的思想,据此制定了一套物种濒危标准(表1).
1994年11月世界自然保护联盟第40次理事会会议正式将经过修订的梅斯-兰德物种濒危等级标准作为新的世界自然保护联盟濒危物种等级标准系统. 最高级别是绝灭,其次是野外绝灭;极危、濒危和易危3个级别统称“受危”;其他依次为依赖保护、接近受危和需予保护,这3个级别也称为低危. 有些则缺乏数据或尚未评估(图1).
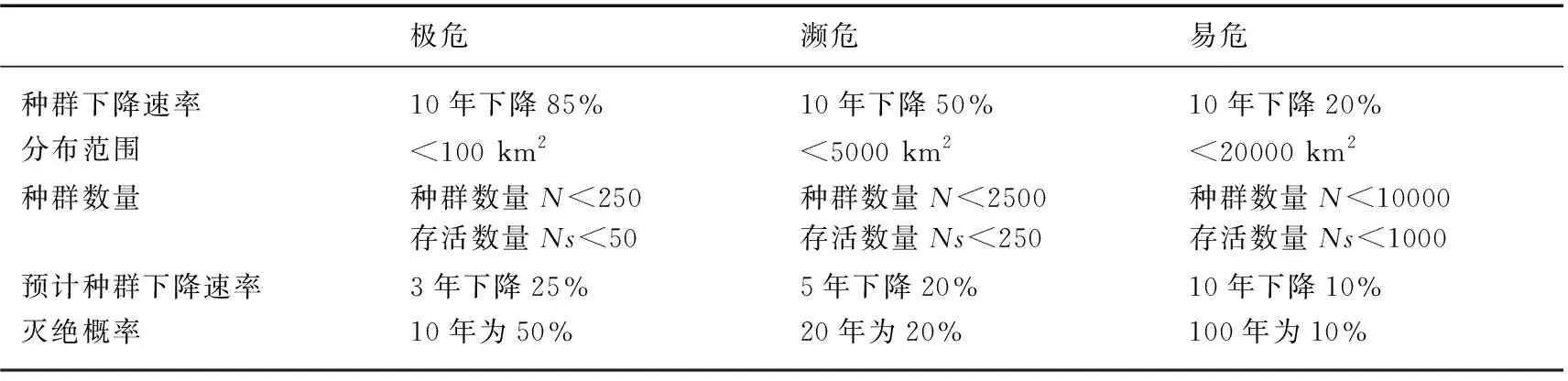
表1 Mace-Lande物种濒危等级标准

图1 世界自然保护联盟物种濒危标准体系Fig.1 World Conservation Union’s standard category of endangered species

3 关于物种濒危机制的理论探讨
为了对濒危物种进行有效拯救,人们必须知晓物种的灭绝机制. 为此,人们提出了最小生存种群、有效种群大小、种群瓶颈、奠基者效应、“阿利”效应、灭绝漩涡等诸多概念.
3.1 最小生存种群
如果能确定某一濒危种群得以生存下去的最低种群数量,对保护实践来说就具有重要意义. Shaffer[5]提出了最小生存种群(minimum viable population,MVP)的概念,指某个物种在野外能够长期生存下去的种群数量下限(或最小生存种群),在此种群规模,物种不会因为自然灾害、统计随机性、环境随机性或遗传随机性而灭绝. 需要指出的是,有些物种虽然在野生环境中灭绝了,但却被人工饲养着.
人们常常用基于计算机模拟的种群生存力分析(population viability analysis, PVA)来估算最小生存种群. 种群生存力分析包括出生-死亡过程模型、矩阵模型、种群随机模型(单一种群、复合种群、空间显示)、以及基于个体的模拟模型等[6].
人们亦以一定时间范围(如100年、500年或1000年)中的生存概率(95%、99%或任何百分比)来判定最小生存种群. 但实际上确定这样的生存概率是非常困难的,因为对一个自然物种的灭绝过程进行超过100年以上的观测研究几乎是不大可能的.
即便如此,一些科学家还是大胆地提出了一些估计数字. 譬如,对陆生脊椎动物而言,MVP值一般是500~1000,如果考虑到近交的效应,许多物种的MVP的估值为数千,譬如,Traill等[7]通过对200多个物种的详细数据分析认为,大部分物种的长期最小生存种群值为3000~5000,平均为4169.
在一定的条件下,根据种群大小可以衡量一个物种的兴衰,因此,如果一个物种分布范围狭窄、仅占据稀有的特殊生境而且种群规模也很小的话,就比一般常见的种类更容易灭绝[8]. 也就是说,一个物种的濒危可根据MVP来进行判断.
但是,现实情况远比上述理论复杂. 譬如,在长江中下游生活的白鱀豚和江豚,虽然食物基本相同,但白鱀豚已经功能性灭绝,而江豚还有约1000头,虽然处于极危状态,这可能与它们不同的生态或行为习性等有关,即它们可能具有不同的最小生存种群,但如果它们的种群降低到各自的阈值之下,可能的“阿利”效应将使其进入灭绝漩涡. 与陆生动物不同,水生动物由于难以直接观察,其最小生存种群更难确认,譬如白鱀豚、江豚、中华鲟等大型水生动物的活动范围跨越数千公里,种群数量的估算均十分困难,迄今为止,无人知晓它们的MVP应该是多大.
事实上,根据MVP对物种濒危的判断并不那么容易,因为有些物种虽然稀有,但可以长期存在下去,等到适宜的环境或机会来临,又会不断壮大起来,而有些所谓的优势物种,也许只要一个关键性限制因子的出现,就会使其快速衰落. 譬如, 20世纪70年代,中华鲟的繁殖群体还有1万多尾,随着20世纪80年代初葛洲坝的修建,其种群迅速衰落,因在葛洲坝下适合其产卵之地寥寥无几,无论中华鲟原来的繁殖群体多大,都难逃绝灭之命. 在这种情况下,所谓的MVP就毫无意义了. 另一个物种——白鲟(图2)比中华鲟更惨,早已不见踪迹.

图2 已经绝灭的世界上最长的淡水鱼——白鲟(Psephurus gladius)(黄宏金等拍摄)Fig.2 Extinct Chinese paddlefish, the longest fish in the world (photo from Huang HJ et al)


图3 已经绝灭的大鳞白鱼[9]Fig.3 Extinct Anabarilius macrolepis[9]

图4 已经绝灭的异龙鲤[14]Fig.4 Extinct Cyprinus yilongensisi[14]

图5 已经绝灭的茶卡高原鳅[15]Fig.5 Extinct Triplophysa cakaensis[15]
3.2 普适性的最小生存种群并不存在
从生态学上来说,种群密度与身体质量是密切相关的,即一定面积中能够栖息的物种的最大种群密度与身体的重量呈现很好的负相关关系,在对数尺度上,两者呈近乎直线的相关关系(图6). 道理很简单,较大的动物需要更多的食物和空间用于栖息、繁殖与生长. 例如一个狮群的领地面积从20到400 km2不等,因此它们的种群数量注定会十分稀少,而一些小型无脊椎动物,如一些昆虫幼虫的活动范围可能局限于产卵地点几厘米范围之内,但密度可能会高得惊人[16].

图6 物种(从病毒到红杉、鲸鱼)的密度与身体质量之间的关系[17]Fig.6 The relationship between density and body mass across species ranging in size from viruses to sequoia trees and whales (data from Damuth 1987)[17]
种群密度与身体质量的这种关系在生理上十分相似的种群中也是成立的,而且无论食性如何,虽然不同食性之间略有差别(图7),即同样的体重,位于不同营养级的哺乳动物其种群密度也呈现出一定的趋势性差异:牧食者>杂食者>肉食者,其实这与它们对应的资源量是相吻合的,即由于能量在营养级间大量损失,高营养级生物的可利用资源量显著减少,所能支撑的种群数量也就显著降低[16].

图7 陆生哺乳动物密度(no/km2)的对数值与平均体重(g)的对数值之间的关系[18]Fig.7 The log of population density (no/km2) plotted against the log of average body mass (g) for terrestrial mammals[18]
因此,不可能存在一个普适性的最小生存种群,不同物种之间可能会有天壤之别. 其实,在生态学意义上的种群规模与身体质量密切相关,大动物的种群数量与小动物不可比拟,很显然,大动物的个体需要大的领地与资源,因此,这样的物种容易濒危或灭绝.
3.3 其它概念
有效种群大小(effective population size,Ne)指一个种群中能将其基因连续传递到下一代的个体平均数,相当于理想状态下的种群大小. 由于种群中许多个体不能生育以及限制个体寻找配偶的社会结构等,均会导致Ne少于实际种群大小(N),而生境退化和片段化等均会引起并加重这些因素对有效种群大小的影响[19]. 或许只有在物种行将灭绝的时刻,才有需要考虑这样的因素.
奠基者效应(founder effect)指当一些个体离开原种群而建立一个新的种群时,新种群的遗传变异性比原来较大种群低的现象. 该概念可追溯至Ernst Mayr和Sewall Wright[20-21]. 大的种群虽然保有更多的遗传变异,但从地史时间尺度来看,新种群恰好能为物种分化带来新的契机.
种群瓶颈(population bottleneck)指当一个种群数量急剧减少、没有携带稀有等位基因个体的生存和繁殖而丢失这些等位基因时,等位基因的减少及杂合度的降低会使种群中个体的适合度降低的现象. 但问题是个体的适合度并不仅仅是由杂合度所决定的,还取决于生存环境. 此外,基因变异也不会歇息.
“阿利”效应(Allee effect):当种群大小和密度跌落到一定水平时,许多动物的社会系统和繁殖系统就会瓦解[22]. 该概念可追溯至20世纪30年代美国生态学家Warder Clyde Allee利用金鱼(goldfish)所做的实验研究,他发现水族箱中有较多个体时,生长更快. 这与密度制约效应是相反的观点,如果考虑群体捕食或群体防御,“阿利”效应不难理解. “阿利”效应也可视为一种正的密度依赖效应,或种群密度与个体适合度(fitness)之间的正相关关系[23].
灭绝漩涡(extinction vortex):小种群的衰退直至灭绝就像一个漩涡,一旦被卷入,物种就不可避免地走向绝灭[24]. 灭绝漩涡包括一系列的模型,保护生物学家、遗传学家和生态学家试图藉此来解析物种灭绝的过程与机理.
MVP、奠基者效应、种群瓶颈等都是所谓的小种群问题,如果某些物种的数量极其稀少,濒临灭绝,就称之为极小种群. 一般认为,小种群或极小种群会出现适合度降低的现象,灭绝风险高. 譬如,欧洲野牛(Bisonbonasus)(图8)在20世纪初差点灭绝了,现在存活的个体都是12头野牛的后裔,它们的遗传变异极低,人们担心会影响它们的繁殖能力[25](表2).

图8 欧洲野牛Fig.8 European bison
这些概念多是一些理论思考,可操作性不强,因此,对保护生物学的实际指导意义不大,有些逻辑也不一定成立. 譬如,从地史时间尺度来看,任何新物种的诞生都源于小种群,因此,问题并不都在于小种群本身.
4 物种生存状态的转换
既然基于种群数量来判断一个物种是否濒危存在诸多问题,可否基于一个物种在生态系统中的生存状态来予以评价?众多物种可能共享一个这样的生态系统(譬如在长江中生活有近400种鱼类),但对每个物种来说,它们的生态位大小不一,在一定历史时期的生存状态可能差异很大(这决定了动物的区域性地理格局),而生态系统的人为扰动(如建坝)对不同物种的生态效应可能也会绝然不同.
在此,笔者借用Scheffer等[26]的生态系统稳态转化概念来描述物种的生存状态转化(Regime shift of species status). 这里,物种的生存状态主要取决于物种对外在环境的生存适合度S和灭绝风险R(图9). 影响到物种生存的外在因素(灭绝外力)有多种多样的形式,譬如,①物种生存空间的完整性(如建坝对鲟鱼和胭脂鱼生殖洄游通道的阻断)、②生态过程的适宜性(如改变了中华鲟产卵的温度过程)、③食物的可得性(如江湖阻隔大大减少了长江干流中白鱀豚和江豚的饵料生物量)、④对人为致死(如江豚的误捕致死、螺旋桨致死等)或自然灾害的抵抗力,等等. 某个灭绝外力越大,相应的物种适合度就越低. 灭绝风险是物种对灭绝外力的遗传、生理、行为和生态等的综合响应,如果数量不足一定水平(类似于“最小生存种群”,但并非一个绝对数量),就可能步入“灭绝漩涡”. 这里的灭绝漩涡指物种所处的一种无法挽救的生存状态,终点就是灭绝.

图9 物种生存状态转换机制概念模型Fig.9 Conceptual model for regime shift of species status
当灭绝外力很小时,物种生存适合度高,处于健康状态,随着灭绝外力的增加,生存适合度降低,物种灭绝风险逐渐增大. 当综合生存适合度降低到B1时,物种的生存状态出现突变,从健康状态进入濒危状态,如果灭绝外力进一步增大就使物种进入灭绝漩涡. 假若我们在物种尚未完全绝灭之前采取有效的措施降低灭绝外力,有时我们必须使综合生存适合度恢复到B2才能使物种从濒危状态恢复到健康状态,即存在时滞(hysteresis)现象. 这里将B1称为物种濒危突变点,B2称为物种拯救临界点. 在这种具有时滞的物种生存状态转化过程中,下降与恢复的轨迹并不完全相同,或者说,同样大小的扰动却无法使其反方向返回原来的状态轨迹,这种现象在生态系统稳态转化中十分常见.
为了定量评估灭绝外力,必须对多种性质不同的胁迫因子进行加权量化,因为任何一个物种都可能会承受多个因子的联合胁迫(如生态位压缩、过度捕捞、食物网破坏、生态过程缺损、生殖洄游通道阻断等). 此外,有些胁迫只有0或1两种状态,如葛洲坝对鲟鱼和胭脂鱼生殖洄游通道的阻断,无坝时胁迫就是0,有坝时胁迫就是1,这几种鱼在坝下均无法形成有效的产卵场,可能因为坝下没有适合其产卵的砾石底质. 此外,即便是同样的胁迫压力,不同物种的灭绝风险R会有所不同.
因此,为了定量计算一个物种的生存状态指数,首先必须确定综合生存适合度f(s),由物种对若干决定性生存因子(determinant survival factor)的适合度(fitness)之乘积所决定:
f(s)=f(s1)f(s2)f(s3)……
(1)
譬如,某种鱼类的决定性生存因子S包括生殖洄游通路的畅通性(s1)、食物可得性(s2)、对人为致死(如误捕致死、螺旋桨致死等)或自然灾害的抵御能力(s3)等. 这些决定性生存因子也表征了相应的灭绝外力. 针对每个决定性生存因子的适合度取值0~1,1表示最大的适合度(理想生存状态),0表示彻底丧失了适合度. 因此,任何一个决定性生存因子的趋零,都会导致该物种的灭绝(木桶效应). 而物种的生存状态指数H(f, R)由f和R联合决定(图9).
当然,笔者还只是提出了一个概念模型,各个决定性生存因子的适合度函数f(s)有待确定,并需要以真实的物种为对象进行验证. 毫无疑问,可用f(s)来评估与预测物种的灭绝风险(譬如,可区分为3~4个风险等级),突变临界点之前的动态过程可作为物种濒危的早期预警信号,物种拯救临界点可作为保护性干预的最低目标. 此外,f(s)还可用于对保护措施进行测试、评估与筛选. 这应该比基于数量、分布范围、种群下降速率对物种濒危进行划分的方法更为合理.
5 迁地保护
即便人们对小种群有这样那样的担忧,但有些迁地保护却十分成功. 任何物种的规模都会经历从小到大的发展过程,小种群也会发展成大种群,大种群也能衰退到小种群甚至绝灭,这主要是取决于生存环境的适宜性,而不是种群数量本身或群体的遗传多样性.
迁地保护(off site conservations)就涉及奠基者效应,一些物种因适宜的生存条件已不复存在,因此种群数量极其稀少以致难以寻找到配偶,需要将其迁出原地进行保护. 迁地保护有很多成功的案例,如天鹅洲故道中的江豚,已自然繁殖出数十头幼豚. 又如,麋鹿(Elaphurusdavidianus)原产于中国长江中下游沼泽地带,在汉朝末年近乎绝种. 到19世纪时,只剩在北京南海子皇家猎苑内一群,后被八国联军捕捉并从此在中国消失,1898年被英国购买并繁殖到255头,1983年部分个体被送回中国, 2011年在江苏盐城大丰湿地的麋鹿接近1800头,2013年在湖北石首市天鹅洲麋鹿保护区的麋鹿超过1000头(图10).

图10 麋鹿Fig.10 Large Chinese deer
迁地保护并不都能成功,生态位的大小亦是重要的因素之一. 一般来说,水生动物的迁地保护比陆生动物更难,如有些种类生活史十分复杂,既需要在海洋中肥育,又需要溯河洄游到江河的淡水环境中去繁殖,迁徙范围达数千公里,中华鲟就是这样一个物种,对中华鲟进行迁地保护就难以想象. 因此,对中华鲟这样的鱼类来说,生殖洄游通道的切断就意味着无可挽回的灭绝.
6 物种的濒危机制与拯救措施
围绕物种的濒危机制,虽然人们提出了很多概念与模型,但很难吻合每一个物种的情形,虽然有些成功,但不乏失败,特别是,珍稀水生生物的濒危与拯救尤其复杂与困难. 下面以中华鲟为例来说明一个物种的濒危机制研究与有效的拯救是多么艰难的一件事(表2).

表2 中华鲟保护大事记*
*主要来源于文献[32-35].
20世纪80年代初,学术界围绕葛洲坝的救鱼展开了广泛的调查、研究与争论(见水电部驻葛洲坝代表处和长江水利委员会编辑的《葛洲坝工程重大技术问题讨论文集》,内部材料,1989年),提出了若干救鱼对象和措施的意见:
1) “中华鲟在葛洲坝下产卵和正常孵化是事实,已在坝下形成了新的产卵场,能在坝下江段依靠天然繁殖保持该物种”. 这一结论主要是依据:①在下游的沙市江段、崇明岛附近共采集到了130尾中华鲟幼鱼,这两批幼鲟是大江截流后的1981年秋出生的;②11月1日至10日,在宜昌的十里红江段捕到了4批铜鱼共149尾,发现有46尾肠管中含有中华鲟鱼卵. 在之后的很多年都能采到少量幼鲟及鱼卵,这是事实,但问题是,只有很小的繁殖规模,根本无法维持原来的自然种群.
2) “葛洲坝枢纽的救鱼对象,主要是中华鲟”,事实证明这并不完全正确,白鲟早就不见了踪影,可能已经灭绝了.
3) “葛洲坝枢纽的救护中华鲟措施,无论从可行性或从合理性来说,都是以人工繁殖放流为佳,建议采用这一措施”. 1984年之后,每年都实施了人工放流,共计向长江中投放了数百万尾中华鲟幼鱼,但存活者似乎寥寥无几,丝毫没有阻挡住中华鲟野生种群衰退的趋势.
最近的研究关注到,三峡蓄水运行后,葛洲坝下中华鲟产卵场江段的水温在10-11月份上升了约2.7℃,导致中华鲟的自然繁殖日期推迟了近1个月. 因此认为,如果通过梯级水库调度将10月22日前的水温降低到18~20℃,可将中华鲟野生种群数量恢复到葛洲坝截流后的水平[27-28]. 且不说能否通过梯级水库调度将水温降低到目标水平,有何依据说中华鲟野生种群数量藉此就可以恢复到葛洲坝截流后的水平?事实上,葛洲坝截留与三峡大坝截留之间,中华鲟繁殖群体的数量已经大幅下降了,其衰落已接近尾声,关于之后的进一步下降是否与三峡大坝蓄水带来的秋季河水增温效应导致的繁殖时间推迟(从而认为抑制了中华鲟的产卵繁殖行为)之间具有真正的因果关系,应该还没有得到有效的论证.
在其它江河修筑水坝不会导致鲟鱼灭绝并不能保证世界上没有例外,在长江上修建葛洲坝对中华鲟看来就是一个例外. 人工放流能拯救其它河流中的鲟鱼,但对长江中的中华鲟来说,似乎也是一个例外. 为中华鲟修建鱼道也许的确无用,也许真不值得花费这个冤枉钱,但问题是在其它措施都不管用时,这也就根本不值去炫耀了. 依据过往经验进行推理所得出的判断并不都会是正确的,这无可非议,只是表述时不要坚信过分. 葛洲坝建成后,坝下中华鲟性腺出现了退化,被认为与游程和流速有关,因其产卵场还需上溯1000 km,那里流速往往在3 m/s甚至达到6~7 m/s[29],这也只是一种推测而已. 其实,其它河流的水坝也有导致鲟鱼性腺退化的,如在伏尔加格勒下游,俄国鲟性腺被吸收的比例达16%,欧洲鳇达15%,在亚速海-黑海区域更加明显[30]. 只是中华鲟性腺退化主要由增温直接引起的说法[28]需要谨慎. 前车之鉴:对通过水温调节来拯救中华鲟亦不可期望太高.
葛洲坝水利枢纽从1969年提出修建、1981年建成、运行至今,已经过去了半个世纪. 建坝前对如何保护中华鲟的问题也进行了详细的论证,建坝后也尽了最大的努力去拯救中华鲟(表2),仅中华鲟研究所从1984年至今就放流了60多次共500多万尾中华鲟,但依然收效甚微. 通过对1996-1998年放流的全长在8~10 cm规格幼鲟的标记回捕试验,估算出这一规格的人工放流个体仅占长江口中华鲟幼鱼群体数量1.71%~3.03%[31].
中华鲟野生种群的灭绝正在进入倒计时. 农业部在2015年发布的《中华鲟拯救行动计划(2015-2030年)》中指出:“长江中华鲟繁殖群体规模已由20世纪70年代的10000余尾下降至目前的不足100尾. 葛洲坝截流至今33年来,中华鲟繁殖群体年均下降速率达到约10%,情况令人担忧. 如不采取有效措施,中华鲟自然种群将迅速走向灭绝”.
7 展望
像任何生命个体一样,物种也是生生死死,据估计,地球上诞生的物种99%以上都灭绝了,物种的平均寿命约为100~1000万年. 就像每个物种的独特性一样,物种灭绝的原因(无论是自然的还是人为的)也是各不相同,有些复杂,有些简单,既有生态的,也有遗传的,等等. 但问题是,地史上的物种灭绝(曾经出现过5次大的灭绝事件)往往可以伴随新物种的大量产生(如寒武纪生命大爆发),而现代灭绝(称之为第6次大灭绝)却不能,因为灭绝速率太快,物种根本来不及适应.
关于现代灭绝,种群数量当然是物种兴衰的重要表征,当它寥寥无几时,人们开始惊呼它“濒危”,但人们往往很难觉察到濒危的早期信号,因为我们并没有经过实践反复检验过的准确的量化模型. 有效的拯救不仅需要人们对濒危机制的透彻解析,譬如,需要了解完成生活史所需的时空(如江湖连通性)完整性、生态过程(如水文过程、温度过程)完整性,等等,还需要构建关于物种生存状态的定量预测模型,并不断在实践中测试与评估,对关键参数反复优化,确定可靠的物种濒危突变点、早期预警信号以及物种拯救临界点等,采取有效措施,及时阻止物种落入灭绝漩涡之中.
无论对物种濒危如何评估,人们对物种保护的努力并未歇息,如建立自然保护区、为野生动物栖息地立法以及设立生态系统红色名录等,其实,这些都与物种的生存状态息息相关. 早在19世纪初,人类就开始了以珍稀濒危野生动植物物种保护为目的的现代自然保护区的建设. 从实践上来说,保护区既有成功的,也有失败的. 在过去的半个世纪,长江的生物多样性面临了很大的危机,但在长江设立的一些水生动物保护区并未起到预期的作用,譬如,几个白鱀豚保护区并未能拯救白鱀豚,几个中华鲟保护区也未能阻挡住中华鲟灭绝的步伐,与陆生动物相比,水生动物的保护任务似乎更为艰巨[36]. 有效的保护实践离不开对物种濒危状态的准确评估以及对未来趋势与突变点的合理预判,否者可能会使物种拯救的宝贵时间丧失殆尽. 另一方面,在很多情况下,人们过分信赖现状去制定对策,却忘记了对历史(特别是自然历史)的谨慎回溯,而也许正确的钥匙就匿藏在那里[37].
