高食性重叠度对同域物种生境分离的影响1)
——以马鹿和狍为例
2020-03-26吕忠海冯源于沿泽张明海张玮琪
吕忠海 冯源 于沿泽 张明海 张玮琪
(东北林业大学,哈尔滨,150040) (海南大学) (黑龙江省林业科学院) (东北林业大学)
同域分布物种是种间竞争研究的理想对象。生态位理论认为,同域分布的相似物种面临食物、水分、空间和营养元素的竞争,会在资源利用的时间、空间和方式等多个维度分化,形成特定的分布格局[1]。同域分布物种的相互干扰、竞争,已经在食肉动物的冲突和共存、大型食草动物的时间生态位分化、部分昆虫对资源利用的时间和空间分化上开展了相关研究,并取得了一些重要进展[2-3]。但由于物种间相互干扰、竞争的表现形式多样,作用机制复杂,仍有许多问题尚未解决[4]。有蹄类动物个体较大,易于观察,是野生动物行为生态学研究的重点。目前对同域分布有蹄类动物的研究,主要集中在食物选择、生境选择的重叠与分化等单方面的研究,尚缺乏同域分布有蹄类动物生境选择及食性协同作用下相互干扰、竞争的研究[1,5-9]。
马鹿(Cervuselaphus)、狍(Capreoluscapreolus)是我国东北地区重要的鹿科经济动物。黑龙江穆棱东北红豆杉国家级自然保护区,位于长白山脉的北端,属于太平岭和乌苏里褶皱带。区内山脉属长白山系,植物资源生长茂盛,野生动物资源丰富,是马鹿与狍等有蹄类动物的重要分布区。而且近年该地区发现有东北虎活动,证明该地区东北虎主要猎物的马鹿和狍种群数量丰富。2013年和2014年1月份,对穆棱东北红豆杉国家级自然保护区内马鹿和狍的生境选择及食性进行了比较研究,从理论上探讨同域分布的马鹿和狍的生境选择及食性的差异,为制定保护区有蹄类动物的保护管理计划提、保护和发展其种群提供必要的基础资料。
1 研究区自然概况
黑龙江穆棱东北红豆杉国家级自然保护区,位于黑龙江省牡丹江市穆棱镇,由穆棱林业局所属的和平林场、龙爪沟林场以及杨木桥林场和共和经营所的部分施业区组成,地理坐标为东经130°0′~130°28′、北纬43°49′~44°6′,东西宽33 km、南北长31 km,总面积约为35 648 hm2。保护区位于长白山脉的北端,属于太平岭和乌苏里褶皱带,地貌以山地为主,属于构造地貌,呈条带状分布。主要有暗棕色森林土、沼泽土、暗色草甸土等土壤;海拨450~900 m。属于典型的温带大陆性季风气候;气温年较差大,年平均气温-2 ℃左右,雨季较短且降水集中,平均降水量514 mm,多在6—8月份;日照时间2 439~2 750 h。
保护区范围内有植物163科472属1 135种,除了东北红豆杉(Taxuscuspidata)外,还分布有黄檗(Phellodendronamurence)、胡桃楸(Juglansmandshurica)、红松(Pinuskoraiensis)、钻天柳(Choseniaarbutifolia)、紫椴(Tiliaamurensis)等珍稀濒危植物以及云杉(Aaliustrinervius)、冷杉(Abiesfabri)、白桦(Betulaplatyphylla)、蒙古栎(Quercusmongolica)、落叶松(Larixgmelinii)等温带针阔混交林的代表性植物。穆棱保护区山地丘陵众多,地势多变,植物群落丰富,结构稳定,为多种野生动物特别是脊椎动物提供充足的食物资源和隐蔽条件。
2 研究方法
2.1 野外数据及样品采集
2013年和2014年1月份,在穆棱东北红豆杉国家级自然保护区内布设样线,马鹿和狍的所有生境类型。在样线调查过程中收集马鹿和狍的新鲜粪便,并记录采集相关生境因子信息。在粪便采集过程中,沿途采集其活动区域内所能见到的每种植物。
生境因子主要记录:(1)植被类型;(2)马鹿食物丰盛度(当年生高度<1.8 m的可食食物枝条数)、狍食物丰盛度(当年生高度<1.5 m的可食食物枝条数);(3)雪深(用直尺测量);(4)坡度(用罗盘测量,坡度划分为>20°、<20°);(5)坡向(用罗盘测定,坡向划分为阳坡、半阴半阳坡、阴坡);(6)坡位(通过目测,人为划分为下坡位、中坡位、上坡位);(7)海拔(用GPS记录);(8)隐蔽度(在选取样方中心位置立1个1.5 m高的杆,10 m外自东南西北方向记录木杆可见高度占总长的比列,并计算其平均值);(9)乔木密度(统计10 m×10 m样方中乔木的数量);(10)灌木密度(统计10 m×10 m样方中灌木的数量);(11)人为干扰距离(用GPS测定)。
2.2 生境因子数据处理
非量化生境因子(植被类型等),统计其各类数据所占百分比,对比分析出马鹿、狍生境因子间的差别;可量化生境因子(海拔等),通过spss软件逐步判别分析法对比分析出马鹿、狍生境因子间的差别。
2.3 食性分析方法
将采集到的粪便样品每份取10粒置于烘箱中60 ℃烘干48 h,粉碎、研磨、过筛。取样品0.5 g置于培养皿中,加体积分数为10%次氯酸钠溶液,消化8 h后制成粪便显微装片,每组粪便样品制片10张。将采集到的的植物样品在烘箱中60 ℃烘干48 h,至恒质量,粉碎、过筛。剩余步骤与粪便样品装片制备相同,每组植物样品制片3张。将粪便样品和植物对照显微装片通过100倍显微镜镜检,每张装片随机选取10个视野,对照植物装片鉴别种类。
通过公式F=100(1-e-D),将统计的每种植物可辨认表皮角质碎片的出现频率(F)转换为每个视野每种植物可辨认角质碎片的平均密度(D),然后将D转换为相对密度(Dr),Dr=(每种植物可辨认的表皮角质碎片密度(D)/各种植物可辨认角质碎片的密度和)×100%。Dr即食物中各种植物的实际比例估计值[10]。

通过Schoener食物重叠指数分析马鹿与狍的食物重叠度[11],Dij=1-0.5(∑|Pik-Pjk|),Dij为马鹿和狍的食物重叠度,Pik、Pjk分别为马鹿与狍共同采食植物在各自取食植物中所占百分比。
3 结果与分析
本次调查共布设样线30条,每条样线5 km,样线间距离大于2 km。共收集马鹿足(卧、粪)迹84个,狍足(卧、粪)迹62个;马鹿粪便48份,狍粪便45份。
3.1 马鹿与狍食性选择对比
根据镜检结果统计(见表1),马鹿共取食植物15种,青楷槭(Acertegmentosum)、东北红豆杉、小楷槭(Acertschonoskii)、木贼(Equisetumhyemale)、青杨(Populuscathayana)、瘤枝卫矛(Euonymusverrucosus)、花楷槭(Acerukurunduense)、黄心卫矛(Euonymusmacropterus)、水曲柳(Fraxinusmandshurica)、紫椴、绣线菊(Spiraeasalicifolia)、东北风毛菊(Saussureamanshurica)、卫矛(Euonymusalatus)、榆(Ulmuspumila)、辽东楤木(Araliaelata);狍共取食植物14种,除东北红豆杉外,其余采食种类与马鹿相同。
由表1可见:马鹿取食植物中相对密度(Dr)大于5%的有8种植物,青楷槭(19.19%)、东北红豆杉(16.64%)、小楷槭(9.71%)、木贼(7.13%)、青杨(7.64%)、瘤枝卫矛(7.17%)、花楷槭(5.23%)、黄心卫矛(5.18%);狍取食植物中相对密度(Dr)大于5%的有8种植物,青楷槭(20.61%)、木贼(18.07%)、青杨(15.18%)、瘤枝卫矛(5.98%)、花楷槭(5.52%)、水曲柳(5.50%)、紫椴(5.30%)。
对比马鹿和狍的取食植物种类,狍除不取食东北红豆杉外,其余植物种类与马鹿采食植物完全相同,二者的食物重叠度达到了84.8%。穆棱林区马鹿冬季取食木本植物种类为13种(乔木7种,灌木6种),这与陈华鹏关于东北林区马鹿冬季采食植物组成木本植物达到98.8%的结论相符合[14];狍取食木本植物种类为12种(乔木6种,灌木6种)。由表2可见,马鹿与狍取食植物多样性指数都比较高,但其营养生态位宽度指数都相对很低。这与东北地区冬季穆棱林区鹿科动物可采食植物种类相对较少相符合。

表1 马鹿与狍食性相对密度及取食序位

表2 马鹿与狍食物多样性及生态位宽度对比
注:食性重叠度84.8%。
3.2 马鹿与狍生境选择对比
统计分析马鹿和狍非量化生境因子(见表3):马鹿主要选择坡度小于20°、阳坡、人为干扰距离大于1 km、针阔混交林的生境,坡位选择相对比较均匀,未见明显的倾向性选择;狍主要选择坡度小于20°,阳坡,中、下坡位,人为干扰距离小于1 km、灌木林的生境。
量化生境因子通过spss软件判别分析结果表明(见表4):马鹿与狍对生境因子的利用差异性显著(H=22.319,df=6,P=0.002)。从判别函数和自变量相关系数看,海拔、隐蔽度、乔木密度以及食物丰富度对判别函数的贡献较大。马鹿主要栖息在海拔、隐蔽度、食物丰富度较高和乔木密度较小的生境中,而狍主要栖息在海拔、隐蔽度、食物丰富度相对较低和乔木密度较高的生境中。
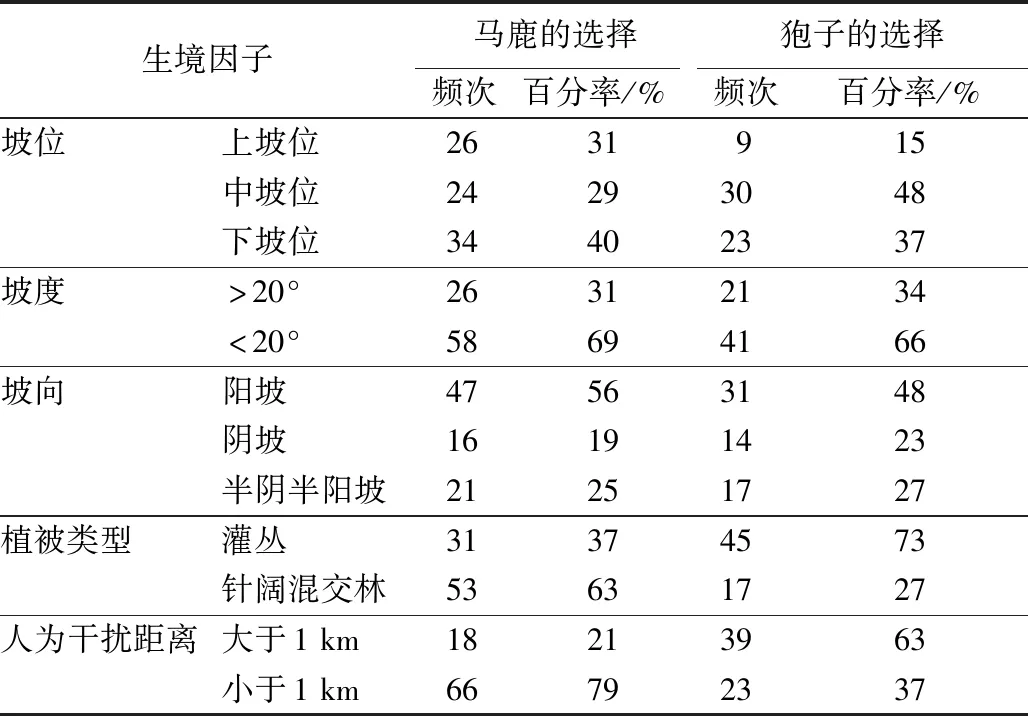
表3 马鹿和狍非量化生境因子分布频次
马鹿与狍生境选择的差异性,主要体现在坡位、人为干扰距离、植被类型、海拔、隐蔽度、乔木密度和食物丰富度7个方面。在东北地区冬季极端条件影响下,可采食植物种类减少,造成同为鹿科动物的马鹿与狍食性产生了很高的重叠(84.8%)。马鹿与狍通过改变资源利用空间分布,从而表现出不冲突行为,达到种间共存。
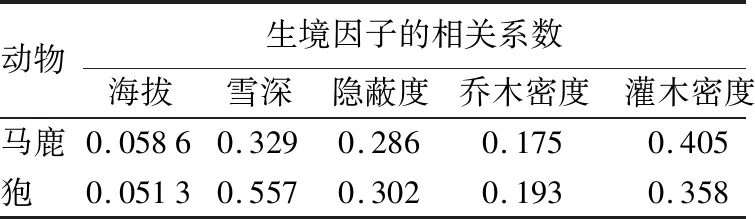
表4 马鹿和狍量化生境因子的判别
4 讨论
营养生态学是研究动物从食物中获取自身所需的各种营养物质,以适应其所处环境的科学[12]。营养对于有蹄类动物而言,是其个体健康和种群发展的重要基础[13]。研究表明,受所处生境的不同及生境因子的不断变化,动物也会通过调整自身趋势策略以适应外界变化[14-15]。由于东北地区冬季积雪覆盖,供马鹿和狍可食用植物种类很少,作为同一地区鹿科动物其食性难免接近。本研究中,马鹿的主要食物为青楷槭(19.19%)、东北红豆杉(16.64%)、小楷槭(9.71%);而狍的主要食物为青楷槭(20.61%)、木贼(18.07%)、青杨(15.18%)。马鹿与狍主要取食植物虽有差别,但二者采食植物种类除东北红豆杉外完全重合,食性重叠度高达84.8%。
有蹄类动物的营养生态位是研究其生境需求的关键所在,对于评价有蹄类动物栖息地、栖息地容纳量以及与其他物种间的生态位竞争与分化有着重要的作用[16]。高斯假说,完全竞争的两个物种不共存[17-18]。生态位理论认为,同域分布的相似物种面临食物、水分、空间和营养元素的竞争,会在资源利用的时间、空间和方式等多个维度分化,形成特定的分布格局[1,19]。马鹿和狍对于生境的选择并非由单一生境因子所决定,而是由多种生境因子综合影响所制约的。本研究中,马鹿与狍受冬季极端条件影响,食物资源受限,食性产生很高的重叠。相互干扰、竞争围绕一个资源中心点展开,优势动物通过攻击或其他方式获得资源的利用权,并干扰和影响其他个体对资源的利用[4,19]。利用几乎相同的食物资源,马鹿与狍种间必然产生竞争。马鹿与狍在生境利用上产生明显分化,马鹿主要栖息于针阔混交林中,狍主要栖息于灌木林中。对不同林型的选择,使马鹿和狍在空间上产生分离,减少相遇的可能性;其次,也是受隐蔽度的影响,针阔混交林隐蔽度较灌木林高。马鹿对隐蔽度要求较高,因而马鹿多选取隐蔽度较高的针阔混交林。马鹿占据了中、上坡位的生境,狍只能选取人为干扰比较强的中、下坡位生境。
根据最优取食理论分析,动物在食物资源量丰富时期,会出现只取食那些最优质量食物的食性特化现象,而当食物资源量匮乏时期,会出现取食更多种食物的食性泛化现象[20-21]。通过对马鹿和狍的食物多样性、营养生态位宽度计算,发现马鹿与狍的食物多样性指数都很高,而生态位宽度却很低。这与东北地区冬季食物资源减少相符合,也证明了马鹿和狍在极端生境条件下,其食性产生了泛化现象。
马鹿和狍作为同域生存物种,其种间关系需要综合考虑生境及营养生态位多方面影响,对其种间竞争关系作出评价。本研究表明,虽然在东北地区冬季极端条件下,马鹿和狍采食植物几乎完全相同,但由于种间竞争关系的影响,其在生境选择及主要食物的偏向性方面已经产生了分化,从而达到了共存的局面,印证了高斯假说。
