海南特有灭绝级植物爪耳木的叶片功能性状
2020-03-25刘咲頔王祝年王清隆叶秀旭
刘咲頔,王祝年, 王清隆, 王 鹏, 叶秀旭
(1.海南大学,海南 海口570100; 2.中国热带农业科学院 热带作物品种资源研究所, 海南 海口 570100)
爪耳木(Lepisanthesunilocularis)隶属无患子科(Sapindaceae)鳞花木属(Lepisanthes)常绿灌木,为国家Ⅲ级重点保护野生植物,海南特有灭绝级植物[1-2]。在消失近80 a后,王清隆等[3-4]在海南省乐东县尖峰镇附近再次发现,但仅有1个居群共5株残存,且是由1株母株通过横走侧根生长出不定芽形成的5株居群,将其移栽至位于海南儋州的南药种质圃(E109°50′9″,N 19°51′9″)中,经过5 a繁育,目前已育成30余株且长势良好。
植物功能性状是指影响植物体存活、生长、繁殖和死亡的一系列植物属性,是植物在生长发育和进化过程中适应外界环境条件的结果[5]。叶片是植物进行光合作用的重要器官,其合成的有机化合物是植物生长发育所消耗能量的主要来源之一。叶片是植物对外界资源条件尤其是光资源变化反应最敏感的器官[6],其功能性状的变化在一定程度上反映植株获取光照资源的能力[7]。叶片功能性状主要分为形态性状、结构性状和生理性状等,其中形态和结构性状与光合速率、水力导度等生理性状密切相关,可一定程度上反映植物与光照、水分及土壤肥力等环境因子的功能关系[8]。
爪耳木具有极耐干旱、耐盐碱的优良特性,是滨海地区较为理想的绿植树种。爪耳木的根可入药,具有一定的经济和药用价值。目前,制约爪耳木种群大小的最大因素是雌雄花不同期且雄花开放极少,导致其自然条件下很难结实,有性生殖受到极大制约。虽然爪耳木极耐干旱,在极端干旱条件下依然不会死亡,但仍将导致其生殖生长几乎停止,其种子在干旱条件下成活率极低。爪耳木是海南特有的灭绝级(EX)珍稀植物,时隔80 a再次被发现,很多特性都尚不明确,研究其叶片功能性状可更好地了解爪耳木的生长习性。鉴于此,采用差异显著性分析、主成分分析及聚类分析等方法,比较新叶与老叶的功能性状差异,探明影响爪耳木生长发育的关键因素及最适生长条件,以期为扩大海南特有灭绝级(EX)濒危植物爪耳木的种群数量提供依据。
1 材料与方法
1.1 材料
1.1.1 叶片 爪耳木当年生新叶和多年生老叶,采自海南儋州南药种质圃(E 109°50′9″,N 19°51′9″)。
1.1.2 试剂 98%浓硫酸、40%氢氧化钠溶液、20 g/L硼酸溶液和0.1 mol/L盐酸溶液,市购。
1.1.3 仪器与设备 AUY220电子分析天平,日本岛津仪器有限公司;EH125ZH酶标仪,奥豪斯公司生产。
1.2 方法
1.2.1 叶片采集 2019年5月17日10:00-11:00在种质圃内选择长势良好的爪耳木植株,采集光照充足无遮挡,无明显病虫害和机械损伤,位于每枝中段生长完全的当年生新叶和多年生老叶各30片,标号(1~60)并称重后置于封口袋中保存。
1.2.2 不同发育阶段爪耳木叶功能性状的差异 将新叶30片和老叶30片各视为1个总体,分别采用单因素方差分析(one-way ANOVA)对其鲜重(WF)、饱水重(WT)、干重(WD)、叶面积(LA)、含水量(LWC)、相对含水量(LRWC)、比叶重(SLW)、全氮含量(LNC)和全磷含量(LPC)进行差异显著性检验(α<0.05)。 鲜重:叶片采下后立即标号并用电子天平称量每片叶的鲜重。饱水重:从封口袋中取出采下的新叶和老叶,分别置入2个盛有清水的保温盒中浸泡,盖上保鲜盒将其放在恒温箱中保存7 h后取出,使用纸巾吸去叶片表面残留的水迹,使用精度为0.01 mg的电子天平称量,记录各叶片的饱水重。干重:将测过饱水重的叶片展开盛于托盘上,105℃杀青20 min,再90℃烘干20 h至恒重后称叶片干重。叶面积:采用画纸称重法测出各新鲜叶片的面积。全氮含量:将干燥处理过的爪耳木叶粉碎,80℃烘干至恒重,过40目筛,称取约0.3 g加入消化管中,每管加入10 mL浓硫酸;待样品充分消解后,采用凯式定氮法[9]测定叶片全氮含量。全磷含量:使用硫酸-过氧化氢消化待测的爪耳木叶干样,使其中存在的各种形态磷转变成正磷酸盐。然后采用钼锑抗显色剂反应测定全磷含量。
根据鲜重、饱水重、干重和叶面积计算比叶重、相对含水量和叶片含水量。
SLW=WF/LA×100%
LRWC=(WF-WD)/(WT-WD)
LWC=(WF-WD)/WF
1.2.3 不同发育阶段爪耳木叶片功能性状因子间的相关性 采用相关性分析法对新叶与老叶间WD、LA、LWC、LRWC、SLW、LNC和LPC性状进行相关性分析。
1.2.4 不同发育阶段爪耳木叶功能性状的主要影响因素 采用因子分析和主成分分析(PCA)探究其性状间的相互影响及性状在不同生长发育过程中的重要性。
1.3 数据统计与分析
采用SAS对数据进行差异显著性分析和主成分分析。
2 结果与分析
2.1 不同发育阶段爪耳木叶片的功能性状
从图1看出,新叶和老叶部分叶片功能性状存在显著性差异。LWC:新叶和老叶的分别为60.21%和52.17%,新叶极显著大于老叶;WD和SLW:老叶和新叶分别为0.046 g、0.037 g和0.017 g、0.013 g,老叶显著大于新叶。LA、LRWC、LNC和LPC:老叶分别为5.670 cm2、72.954%、16.872 g/kg和1.173 g/kg,新叶分别为5.125 cm2、79.807%、16.467 g/kg和1.272 g/kg,各项指标老叶与新叶差异不显著。可见,老叶与新叶的差异主要体现在叶面积、含水量及干物质积累量等方面,叶片的氮、磷含量方面并没有显著差异。说明,爪耳木新叶和老叶代谢的速率相近。
注:图中不同小写字母表示处理间差异显著(P<0.05)。
Note: Different and lowercase letters in the figure indicate significance of difference atP<0.05 level respectively among different treatments.
图1不同发育阶段爪耳木叶片的功能性状
Fig.1 Functional traits ofL.unilocularisleaves at different growth stages
2.2 不同发育阶段爪耳木叶片功能性状因子间的相关性
从表1看出,WD与LWC、LRWC和LPC呈极显著负相关,相关系数分别为-0.568、-0.442和-0.560;LWC与LRWC、LA(r=0.800和0.181),WD与LA、SLW(r=0.614和0.819)均呈极显著正相关;LWC与LPC(r=0.288)呈显著正相关;LRWC与LNC(r=-0.316),LA与LPC(r=-0.305),SLW与LPC(r=-0.634)均呈显著负相关。
对比各性状公因子1和公因子2的绝对值(表2)可知,WD、SLW和LPC受公因子1的影响较大,其公因子的绝对值分别为0.312、0.330和0.279;而LWC、LRWC和LNC受公因子2的影响较大,其公因子的绝对值分别为0.434、0.441和0.238。

表1 爪耳木叶功能性状间的相关性
注:*和**分别表示相关性显著(P<0.05)和极显著(P<0.01)。
Note:* and ** indicate significance of difference atP<0.05 andP<0.01 level respectively.
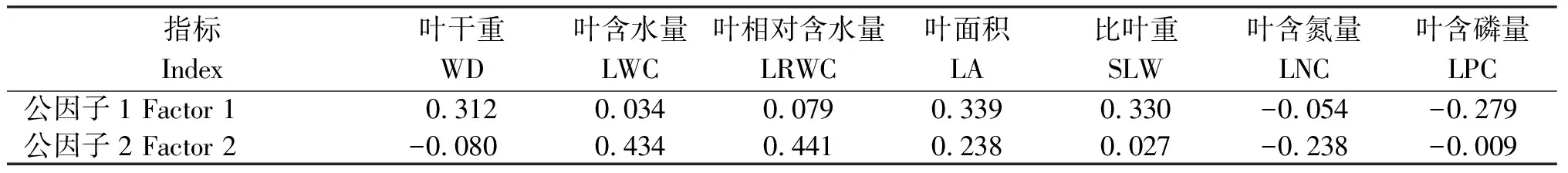
表2 爪耳木叶片功能性状的公因子载荷矩阵
2.3 不同发育阶段爪耳木叶功能性状的主要影响因素
由表3可知,主成分1和主成分2的特征值分别为3.140和1.684(>1),主成分1的贡献率为44.86%,主成分2的贡献率为24.06%,二者的累积贡献率为68.92%。其中,主成分1中以WD、LWC、SLW和LPC所占比重较大,其在主成分1中所占的系数绝对值分别为0.533、-0.393、0.451和-0.396;主成分2中,LWC、LRWC和LA所占比重较大,其在主成分2中所占的系数绝对值分别为0.486、0.527和0.506。说明,WD、LWC、SLW和LPC是主成分1中比较重要的性状;LWC、LRWC和LA是主成分2中比较重要的性状。
表3爪耳木叶片功能性状各主成分的特征值与贡献率
Table 3 Eigenvalue and contribution rate of different principal components of functional traits ofL.unilocularisleaves
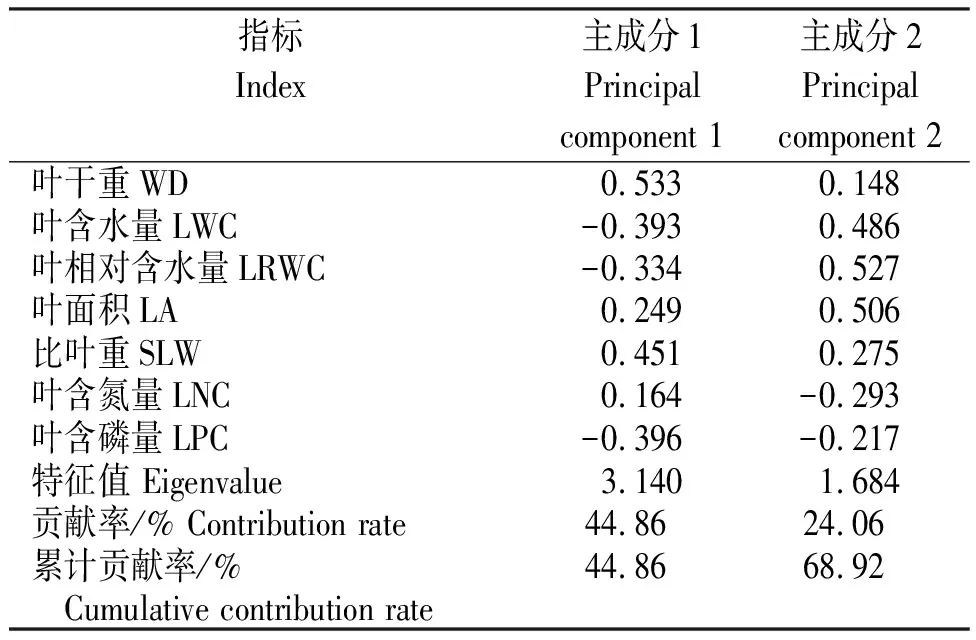
指标Index主成分1Principalcomponent 1主成分2Principal component 2叶干重WD0.5330.148叶含水量LWC-0.3930.486叶相对含水量LRWC-0.3340.527叶面积LA0.2490.506比叶重SLW0.4510.275叶含氮量LNC0.164-0.293叶含磷量LPC-0.396-0.217特征值 Eigenvalue3.1401.684贡献率/% Contribution rate44.8624.06累计贡献率/% 44.8668.92 Cumulative contribution rate
根据各样本的主成分1和主成分2的得分聚类,以主成分1为y轴,主成分2为x轴的散点-聚类(图2)显示,各样本大致聚成2个主要的集团,一个绝大多数由新叶组成,另一个绝大多数是老叶组成,其在坐标轴上的差异主要体现在纵坐标上,即其主成分1得分差异较大。而主成分1中WD、LWC、SLW和LNC所占比重较大,因此,可以判断爪耳木不同生长时期叶片功能性状主要受WD、LWC、SLW和LNC等4个性状的影响。
注:编号1~30为新叶,31~60为老叶。
Note: 1~30 and 31~60 indicates the new leaves and perennial old leaves respectively.
图2爪耳木老叶和新叶叶功能性状PCA分析的散点-聚类
Fig.2 PCA scatter-cluster of functional traits of the old leaves and new leaves ofL.unilocularis
3 结论与讨论
叶片是植物进行光合作用的主要器官,其功能性状能较好地反映植物当前生长发育阶段的一些特性,如所需的营养元素等[10]。叶面积是衡量植物光合能力的重要指标[11],比叶重可作为衡量植物相对生长速率的指标[12]。植物的叶片功能性状可以反映其生长地区的环境情况,其改变可揭露植物生境的演替[13-14]。
叶片含磷量在主成分1中有比较重要的影响。有研究表明,随着生长时期的变化,老叶的磷含量低于新生叶片,原因在于植物叶片获得的营养更多地运输到其他器官用于生根、开花及结实等[15]。爪耳木没有产生这种情况可能是因为其特殊的濒危机制,爪耳木多数是由1株母株通过横走侧根形成的不定芽形成子株,其根部呈网状且扎根非常浅,生长也十分缓慢,自然生长条件下极少结实,如此可能导致其新叶与老叶氮磷含量之间的变化无显著性差异。而老叶与新叶间的差异可能更多体现在干物质的积累量上,这些可从干重和叶面积看出。
叶片氮含量可以说明植物光合能力的高低[16-17];有研究认为,叶片氮含量与光合速率呈显著正相关[18];另有研究认为,叶片氮含量与光合速率呈负相关甚至不相关[19]。研究结果表明,叶干重、叶含水量、比叶重和叶含氮量等4个性状是影响爪耳木叶生长发育的重要因素,今后可以从这些因素入手研究爪耳木的叶片功能性状与其生长发育之间的关系。
研究结果表明,叶片氮含量的增加或减少与其不同生长时期无显著相关性(r=0.560),但并不能说明氮含量在爪耳木叶片生长发育过程中不重要。有研究表明,在与爪耳木同属无患子科龙眼的生产中,发生“冲梢”的个体叶片氮含量显著高于正常开花的个体[20]。叶片氮含量是植株生长的重要影响因素,可能是由于海南位于热带地区,光照强度比较高,新叶和老叶的光合速率趋于1个稳定值,导致叶片氮含量并无明显的上升或下降。后期可对不同生长时期爪耳木叶片的光合特性等进行深入研究。
该研究结果表明,叶含水量是影响爪耳木生长发育的重要因素,也说明水分是影响爪耳木营养生长和生殖生长的关键因素之一。保护爪耳木可以从水分的角度切入,结合氮元素多寡对爪耳木生殖生长的影响,探究爪耳木对环境元素的需求,从而更好地了解其濒危机制,进而采取更多有效手段对其进行保护。
