水稻温敏雄性不育突变体mh86s的鉴定及基因克隆
2020-03-23杨绍华刘华清王锋
杨绍华 刘华清 王锋

摘 要:光/溫敏核雄性不育基因在水稻杂种优势利用中发挥着重要作用。发掘光/温敏核雄性不育突变体及其基因有助于更好地了解水稻中光/温敏核雄性不育的机制并丰富遗传资源。本研究在籼稻材料明恢86的转基因后代中鉴定了1个雄性不育突变体,该突变体在正常生长季表现为雄性不育,花药较野生型小,淡白色,镜检无花粉,孕穗期低温处理后育性恢复,为1个温敏雄性不育突变体,命名为
mh86s。利用突变体mh86s分别和其野生亲本明恢86及籼稻材料93-11杂交,构建F2遗传群体,遗传分析表明雄性不育性状由1对隐性核基因控制。采用图位克隆技术将该温敏雄性不育基因定位在第2染色体分子标记Indel7.38和SNP3之间49.8 kb的物理区间内。在该定位区间内包含已克隆的温敏雄性不育基因tms5。测序结果表明,突变体mh86s中tms5基因和野生型相比,第2外显子1个碱基发生了替换(G-T),导致推测的161位氨基酸由甘氨酸变为缬氨酸。利用突变体mh86s和携带tms5基因的温敏不育系HD9802S杂交,F1表现为温敏雄性不育。据此推断突变体mh86s中的温敏雄性不育基因为tms5的新等位基因,命名为tms5 mh86。研究结果丰富了tms5基因资源,有助于解析tms5基因结构与功能,相应的分子标记亦可用于tms5 mh86的标记辅助选择。
关键词:温敏雄性核不育;基因定位;水稻
中图分类号:S511 文献标志码:A 文章编号:0253-2301(2020)01-0001-05
DOI: 10.13651/j.cnki.fjnykj.2020.01.001
Identification and Gene Cloning of the Thermo sensitive Male Sterile Mutant mh86s in Rice
YANG Shao hua, LIU Hua qing, WANG Feng
(Fujian Provincial Key Laboratory of Genetic Engineering for Agriculture, Biotechnology Institute of
Fujian Academy of Agricultural Sciences, Fuzhou, Fujian 350003, China)
Abstract: Photo or thermo senstive genic male sterile gene (PGMS or TGMS) has played an important role in the utilization of heterosis in rice. The identification of PGMS or TGMS mutants and genes could help to better understand the mechanism of PGMS or TGMS in rice and enrich the genetic resources. In this study, a male sterile mutant was identified from the transgenic offspring of Minghui 86, an indica rice material. The mutant showed male sterility in the normal growth season, the anthers were smaller than those of the wild type, pale white, and no pollen was detected by microscopy. The fertility could be recovered after the low temperature treatment at booting stage, and the mutant was a thermo sensitive male sterile mutant, which was named mh86s. The mutant mh86s was hybridized with their wild parent Minghui 86 and the indica rice material 93-11, respectively, thus to construct the F2 genetical population. And the genetic analysis showed that the male sterile character was controlled by a pair of recessive genic gene. By using the map based cloning, the gene of thermo sensitive male sterility was located in the 49.8 kb physical interval between the molecular markers Indel7.38 and SNP3 of the chromosome 2, and the cloned thermo sensitive male sterility gene tms5 was included in the mapping interval. The sequencing analysis revealed that there was a single base substitution (G to T) in the second exon of tms5 gene in the mutant mh86s compared to the wild type, which led the presumed amino acid was changed from glycine to valine at the 161th position. F1 was characterized by thermo sensitive male sterility by the hybridization of the mutant mh86s and the thermo sensitive male sterile line HD9802S carrying
tms5 gene. Therefore, it was inferred that the gene of thermo sensitive male sterility in the mutant mh86s was the new allele of
tms5, which was named tms5 mh86. The results of this study enriched the tms5 gene resources and contributed to the analysis of tms5 gene structure and function, and the corresponding molecular markers could also be used in the marker assisted selection of tms5 mh86.
Key words: Thermo senstive genic male sterility; Gene mapping; Rice
水稻是一种以自交为主的常异交作物,雄性不育在水稻杂种优势利用中发挥着重要作用,是一种重要的遗传资源。因此发掘水稻雄性不育突变体,充分阐明雄配子发育机制有着重要的理论意义及应用价值。到目前为止,发现并鉴定了大量的水稻雄性不育突变体及其控制基因[1],其中有些突变体为环境敏感型,在不同条件下表现为雄性不育或可育。典型的有受光周期或温度调控的光敏/温敏雄性核不育基因
tms5[2]、pms1[3]、pms3[4]、tms10[5]和csa[6],以及近年来报道的湿度敏感型雄性核不育基因OsOSC12[7]和hms1[8]。
tms5基因编码的RNA酶Zs1,降解3个泛素核糖体L40融合蛋白基因(Ub40)的mRNA,在高温条件下,Ub40基因表达量增加,而突变体核酶功能缺失, Ub40 的mRNA不能降解而积累,导致花粉不育。在低温条件下Ub40基因表达量较低,不影响花粉发育,表现为育性正常[2]。pms1基因编码长链非编码RNA PMS1T,是microRNA2188的靶基因,在miRNA2188的作用下产生21 nt的phasiRNA。Pms1在miRNA靶位点附近的1个SNP造成phasiRNA积累程度的差异,长日照条件下,phasiRNA大量积累导致雄性不育,而短日条件下phasiRNA积累较少,育性正常[3]。pms3是最早发现的光敏雄性核不育突变体农垦58S中鉴定的,
pms3基因编码长链非编码RNA,农垦58S中该基因有1个C突变为G,导致长链非编码RNA甲基化程度发生改变,长日条件降低了长链非编码RNA表达量,从而使得花药发育紊乱而雄性不育[4]。csa基因为MYB转录因子,调控水稻花药中糖的分配,该基因突变后水稻在短日照下表现为雄性不育,而长日照时育性恢复[6]。水稻tms10
突变体表现为高温雄性不育,低温雄性可育的表型,育性转换临界点温度为22~24℃。TMS10编码1个亮氨酸受体激酶,高温条件下TMS10激酶活性在水稻花药绒毡层的降解过程中起重要作用,而在tms10突变体中绒毡层不能正常降解导致花药外壁发育异常[5]。湿度敏感型雄性不育基因OsOSC12为三萜合酶(OsOSC12/OsPTS1)催化2,3 环氧鲨烯产生1种二环三萜化合物禾谷绒毡醇,突变基因导致花粉中禾谷绒毡醇明显减少,使得突变变得更容易脱水而失去活性,测试表明在湿度低于60%的环境中突变体表现为不育,而在湿度高于80%的环境中则表现为完全可育,是一种新型的湿度敏感型雄性核不育材料[7]。hms1基因编码1个β 酮脂酰辅酶A合酶,影响花粉壁中长链脂肪酸的合成,该基因突变后导致突变体花粉壁油层缺陷,使得花粉保水能力下降,在低湿条件迅速皱缩而失去活性,从而导致雄性不育[8]。
由于环境敏感型水稻雄性不育能通过控制环境条件在可育和雄性不育之间转换,因此可以利用其在可育状态下正常繁殖,在雄性不育状态下生产杂交种,能规避三系杂交水稻中恢保关系的制约,在杂种优势利用中发挥重大的作用。发掘和鉴定更多的环境敏感型水稻雄性不育突变体及其控制基因有着重要的应用价值及理论意义。本研究在籼稻品种明恢86的组织培养后代中鉴定了1个温敏雄性核不育突变体明恢86S,对突变基因进行了精细定位,序列分析及等位分析推测突变基因是温敏雄性核不育基因tms5的新等位基因。
1 材料与方法
1.1试验材料
温敏雄性不育突变体明恢86S为本实验室在籼稻品种明恢86组培后代中发现并鉴定。明恢86及杂交亲本籼稻品种93-11,两系不育系HD9802S均为本实验室保存品种。
1.2表型鉴定
抽穗期取即将开花的小穗,解剖镜下观察花药形态特征。花粉育性鉴定采用1% I2 KI染液染色,显微镜下观察并拍照。突变体抽穗育性鉴定为雄性不育后割兜再生,再生株减数分裂期时在22℃低温冷水池处理7 d,然后移栽自然环境,抽穗时鉴定花粉育性。
1.3遗传分析及基因定位
将突变体分别与其野生型明恢86及籼稻品种93-11杂交。F1自交后获得F2。在F2群体抽穗时按单株调查花粉育性并统计分离比。利用突变体和93-11杂交的F2群體中的雄性不育株作为定位群体。从实验室构建的电子图谱中每条染色体选择均匀分布的6个SSR分子标记分析22个不育单株[9],显著偏离孟德尔分离比的分子标记为连锁标记,在连锁标记附近根据93-11和本实验室明恢86重测序序列差异开发Indel和SNP标记进行连锁关系验证并进行基因定位。Indel标记利用3%琼脂糖或8%聚丙烯酰胺凝胶电泳分析。SNP标记采用等位特异性扩增[10],先利用外围引物扩增出靶序列,然后再分别利用位点特异的内部引物和一条相应的外部引物扩增,通过扩增产物的有无判断基因型。
1.4 候選基因分析
采用水稻基因组预测项目网站的基因注释信息(rice genome annotation project,http://rice.plantbiology.msu.edu/)进行基因预测。候选基因通过分段PCR扩增,测序比较突变体和野生型序列差异。
2 结果与分析
2.1 突变体表型特征
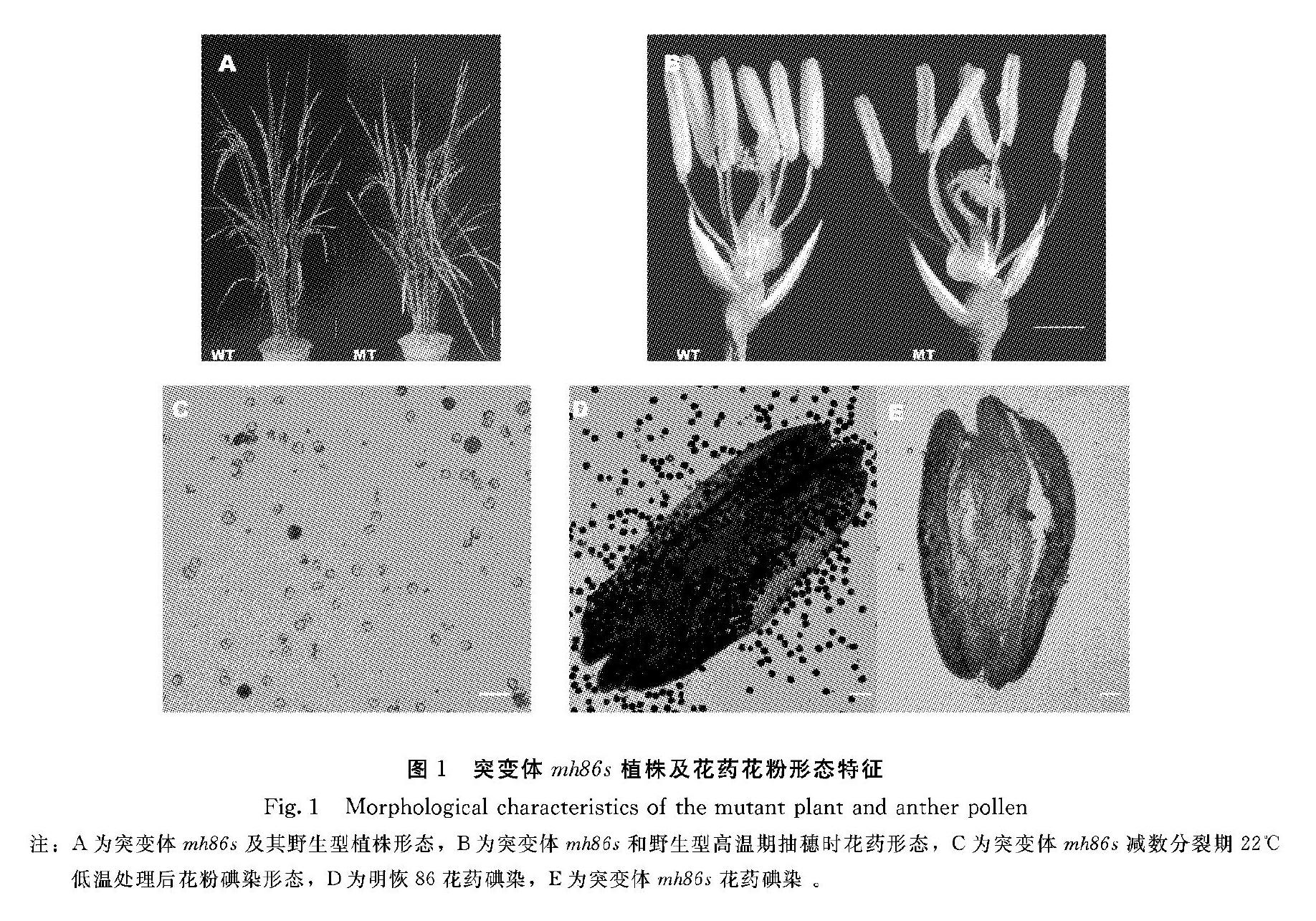
在营养生长期,突变体和野生型表型无明显变化,在福州寿山基地夏季8月高温抽穗扬花时突变体穗不能完全抽出(图1A),花药呈淡黄至白色,偏小(图1B),碘染显示无花粉(图1D)。突变体减数分裂期22℃低温冷水池处理7 d后,待再生穗抽穗时观察发现花药变黄,碘染后显微观察发现大部分花粉为圆败或典败类型,少量为正常黑染类型(图1C)。表明突变体经低温冷水处理后花粉育性得到部分恢复,为温度敏感型雄性不育。因此将突变体命名为明恢86S(minghui86S,mh86s)。
2.2 突变体遗传分析及基因精细定位
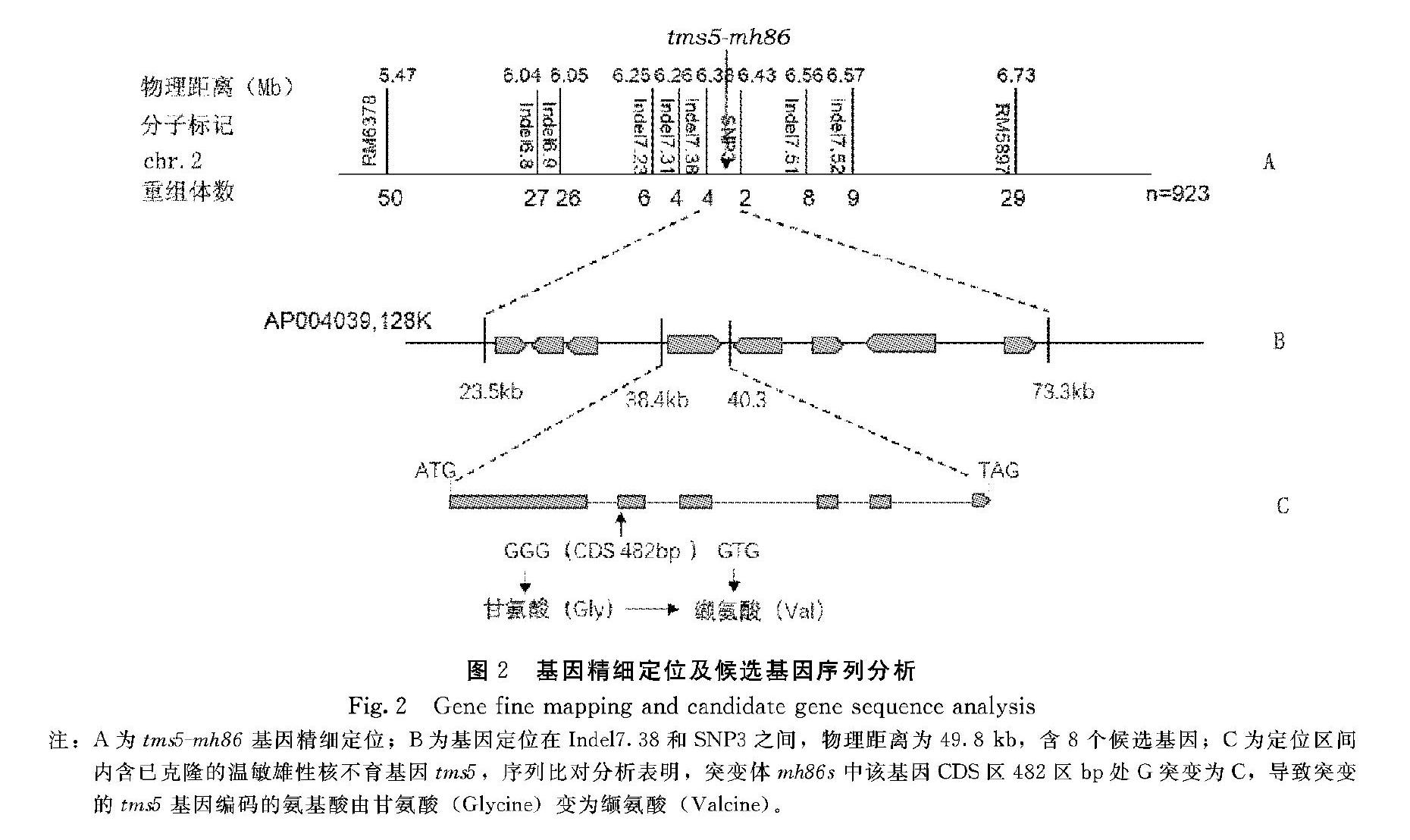
利用突变体mh86s和其野生型杂交,构建F2群体,在福州寿山基地8月抽穗时调查育性,在143株群体中可育株和不育株分别为108和35株,卡方检验分离比符合3∶1(χ2= 0.02<χ20.05,1=3.84),表明突变体mh86s的雄性不育受单一隐性基因控制。利用突变体mh86s和93-11杂交构建的F2群体中的雄性不育株用于连锁分析及基因定位,筛选到位于第2号染色体上的标记RM5897和RM6378和目的基因基因连锁。进一步,将上述2个标记分析172个不育单株,分别检测到9株和6株不同的重组体。上述结果表明突变基因位于两分子标记之间。为精细定位该突变基因,扩大群体到923株,先利用RM5897和RM6378筛选出重组体,再利用新开发的Indel和SNP分子标记对重组体进行基因型鉴定,最终将目的基因定位在水稻第2号染色体分子标记Indel7.38和SNP3之间49.8 kb的物理区间。基因定位用到的分子标记列于表1。
2.3 候选基因分析
根据水稻基因组预测项目网站的基因注释信息可知,精细定位区间内包含8个基因(图2B),其中1个为已克隆的温敏雄性不育基因tms5。因此通过PCR扩增后测序比较突变体mh86s及其野生型品种明恢86的基因组序列。序列比较发现在第2外显子的第2个,即基因CDS区距起始密码ATG下游482 bp处,碱基G替换为T,导致编码第161位氨基酸由甘氨酸变为缬氨酸(图2C)。为验证该突变是否导致mh86s的温敏雄性核不育,以突变体mh86s为母本,含tms5
基因的水稻品种HD9802S为父本杂交,F1在福州8月中旬抽穗时表现为雄性不育,割兜后再生,10月中旬抽穗育性恢复。上述结果表明本研究鉴定的突变位点导致了突变体mh86s
的温敏雄性不育,为tms5的新等位基因,命名为tms5 mh86。
3 讨论与结论
tms5基因是我国水稻两系不育系中广泛使用的光温敏核不育基因资源[4,11-12],突变基因编码一个截短的RNA酶Zs1,导致酶功能缺失,不能正常降解导致雄性不育的UBL40。本研究鉴定的突变基因经等位性测试,表明突变基因为tms5的1个新的等位突变。tms5基因为生产上广泛使用的温敏雄性不育基因,具有重要的应用价值,为探索其作用机制或创制优良的两系不育系,众多研究者利用基因编辑技术创制了大量的tms5基因的等位突变,但这些突变均为提前终止或移码突变等功能缺失突变。而本研究鉴定的等位突变仅存在1个氨基酸的替换,导致单个氨基酸的改变,表明该位点是tms5基因不可或缺的重要的功能位点。但该位点如何影响基因功能还有待进一步研究。
温敏雄性不育起点温度是水稻两系不育系的一个重要参数,较低的起点温度有利于保证杂交种的生产纯度,但过低的起点温度又不利于不育系的自我繁殖。大量研究及实践表明,起点温度和遗传背景密切相关,不同遗传背景起点温度差异很大。突变体mh86s在22℃冷水处理7 d后育性部分恢复,仅极少量结实,表明突变体mh86s转育起点温度较低,可以用于培育两系不育系。研究中发现,定位群体中的不育株秋季低温再生穗结实率差异较大,少部分能正常结实,表明群体中不同个体不育起点温度差异较大,和前人结果一致。因此利用tms5 mh86等位基因有望能培育出具有不同起点温度的两系不育系。本研究鉴定新的tms5等位突变基因丰富了基因资源,有利于对水稻两系不育系不育起点温度的调控,研究中所开发的紧密连锁的分子标记也可利用与分子标记辅助选择。
参考文献:
[1]GUO J X,LIU Y G.Molecular control of male reproductive development and pollen fertility in rice[J].J Integr Plant Biol,2012,54(12):967-978.
[2]ZHOU H,ZHOU M,YANG Y,et al.RNase ZS1 processes UbL40 mRNAs and controls thermosensitive genic male sterility in rice[J].Nature Communications,2014,5:4884.
[3]FAN Y,YANG J,MATHIONI S M,et al.PMS1T,producing phased small interfering RNAs,regulates photoperiod sensitive male sterility in rice[J].Proceedings of the National Academy of Sciences,2016,113(52):15144-15149.
[4]DING J,LU Q,OUYANG Y,et al.A long noncoding RNA regulates photoperiod sensitive male sterility,an essential component of hybrid rice[J].Proceedings of the National Academy of Sciences,2012,109(7):2654-2659.
[5]YU J,HAN J,KIM Y,et al.Two rice receptor like kinases maintain male fertility under changing temperatures[J].Proceedings of the National Academy of Sciences,2017,114(46):12327-12332.
[6]ZHANG H,XU C,HE Y,et al.Mutation in CSA creates a new photoperiod sensitive genic male sterile line applicable for hybrid rice seed production[J].Proceedings of the National Academy of Sciences, 2013,110(1):76-81.
[7]XUE X,ZHOU Y,WANG X,et al.Deficiency of a triterpene pathway results in humidity sensitive genic male sterility in rice[J].Nature Communications,2018,9(1):604-613.
[8]CHEN H Q,ZHANG Z G,NI E D,et al.HMS1 interacts with HMS1I to regulate very long chain fatty acid biosynthesis and the humidity sensitive genic male sterility in rice(Oryza sativa)[J].New Phytol,2019:doi:10.1111/nph.16288.
[9]黃燕娟,华亚鹏,刘华清,等.三个基于水稻明恢86 的SSR电子遗传图谱的构建[J].分子植物育种,2008,6(4):655-663.
[10]NEWTON CR,GRAHAM A,HEPTINSTALL LE,et al.Analysis of any point mutation in DNA:The amplification refractory mutation system (ARMS)[J].Nucleic Acids Res,1989,17:2503-2516.
[11]范优荣,曹晓风,张启发.光温敏雄性不育水稻的研究进展[J].科学通报,2016,61(35):3822-3832.
[12]斯华敏,付亚萍,刘文真,等,水稻光温敏雄性核不育系的系谱分析[J] .作物学报,2012, 38(3):394-407.
(责任编辑:柯文辉)
