基于16S rRNA基因分析虫草寄主蝠蛾的系统进化关系
2020-03-21彭树英万胜豪汪承巧韩蒙蒙汪雨晨耿雪侠张海军
彭树英,李 俊,万胜豪,汪 敏,汪 玲,汪承巧,韩蒙蒙,汪雨晨,耿雪侠,张海军①
(淮北师范大学 生命科学学院,资源植物生物学安徽省重点实验室,安徽 淮北235000)
0 引言
冬虫夏草(Ophiocordyceps sinensis)为麦角菌科(Clavicipitaceae)冬虫夏草菌(Ophiocordyceps sinensis(Berk.)G.H.Sung et al.(Cordyceps sinensis Berk.))寄生在蝠蛾属(Hepialus)昆虫幼虫上长出的子座及幼虫尸体所形成的虫菌复合体[1~3]. 感染此种真菌的昆虫幼虫在冬天形成致密的菌核即“冬虫”,至次年春末夏初之际虫体头部长出子座破土而出,即“夏草”[1-3]. 冬虫夏草是一种极为珍贵的中药材,具有补肺益肾[4]、止咳化痰、调节免疫力[5]、降血糖降血脂[6-7]、抗肿瘤[8]、抗氧化[9]和抗衰老[10]等多种功效. 冬虫夏草寄主昆虫的地理分布虽较独特,但有其特定规律,主要分布于青海、西藏、四川、云南和甘肃五省境内海拔3 000 m以上的青藏高原高寒草甸区[11]. 冬虫夏草菌主要寄主昆虫为蝠蛾属昆虫,根据传统形态学分析,中国境内记录的蝠蛾属昆虫累计达到六十多种,据推测,未发现的种类可能会更多[12-13]. 1968年,Viette[14]以雄性生殖器抱器瓣底部明显的突起为依据,在蝙蝠蛾科下建立钩蝠蛾属(Thitarodes). 随后,国外学者[15~17]将大部分蝠蛾种类归于此属. 由于昆虫形态变化的可塑性及趋同进化导致传统分类法不够准确[18],而且对冬虫夏草寄主的研究也多侧重于形态特征、生物学特性分析,或蝠蛾属昆虫的种类和地理分布的研究[19~25],对我国不同产地冬虫夏草寄主昆虫之间的分子遗传进化关系的研究报道也较少,这将直接影响冬虫夏草种质资源的保护和有效利用[12].
线粒体DNA(mtDNA)为双链共价闭合环状DNA,具有结构简单、无组织特异性、严格遵守母系遗传、不易发生重组、进化速率快等特点[11]. mtDNA是研究动物遗传多样性和分子进化的有效标记,已广泛用于昆虫分类和分子系统学的研究. 一些学者尝试将形态分类方法和分子生物学方法结合,以弥补传统方法的不足. 陈永久等[26]曾利用线粒体细胞色素b 基因(Cytochrome b,Cyt b)序列首次分析5 种冬虫夏草寄主蝠蛾的分子进化关系. 程舟等[12]通过对冬虫夏草寄主蝠蛾的线粒体Cyt b基因片段的序列分析,构建分子系统树,分析我国冬虫夏草寄主蝠蛾种属的系统进化关系和地域分布格局. 邹志文等[13,18]以蝠蛾雄性生殖器抱器瓣结构特征为依据,修订现行中国蝠蛾属昆虫分类系统,将中国蝠蛾属的62个种分别归入拟蝠蛾属(Parahepialus)、蝠蛾属(Hepialus)、钩蝠蛾属(Thitarodes)及无钩蝠蛾属(Ahamus)等4个属中,同时利用26 种原蝠蛾属种类Cyt b基因片段构建系统进化树,证明将蝠蛾雄性抱器瓣特征作为分属依据是合理可信的. 陈抒云等[27]对冬虫夏草和其它虫草等物种的寄主昆虫分别通过COⅠ基因和Cyt b 基因进行条形码分析,表明COⅠ序列更适用于虫草寄主昆虫的DNA 条形码研究. 近几年,一些学者为深入研究虫草寄主蝠蛾的分子进化及其分类,已相继测定多条虫草寄主蝠蛾mtDNA全基因组序列[11,18,28~31].
作为分子标记,研究动物进化的mtDNA应用最多的是Cyt b,细胞色素氧化酶亚基Ⅰ(COⅠ),NADH脱氢酶亚基(ND4),12S rRNA基因,A+T富集区,腺苷三磷酸酶(ATP6或ATP8)等[32]. 16S rRNA基因亦是线粒体基因组中研究较多的基因,既含有保守序列,又含有与进化距离相适应的可变序列,被称为进化分子钟[32]. 16S rRNA基因用于评价生物的遗传多样性和系统发生关系的报道很多[33~38],但在冬虫夏草寄主蝠蛾方面的研究尚未见报道. 本研究运用线粒体16S rRNA基因序列对不同虫草寄主蝠蛾的进化关系进行分析,以期为进一步的遗传系统进化分析、种质鉴定和资源保护奠定理论依据.
1 材料和方法
1.1 基因序列来源
目前已登录至GenBank的蝠蛾科线粒体基因组全序列共有11条记录,其中7条为虫草寄主蝠蛾(其中6种蝠蛾基因序列有两个序列号),亚香棒虫草寄主蝠蛾1条,其他蝠蛾3条(其中2种蝠蛾基因序列有两个序列号),从GenBank中获取并下载蝠蛾科、其他科蛾类及果蝇(外群)线粒体16S rRNA和COⅠ基因序列(表1).
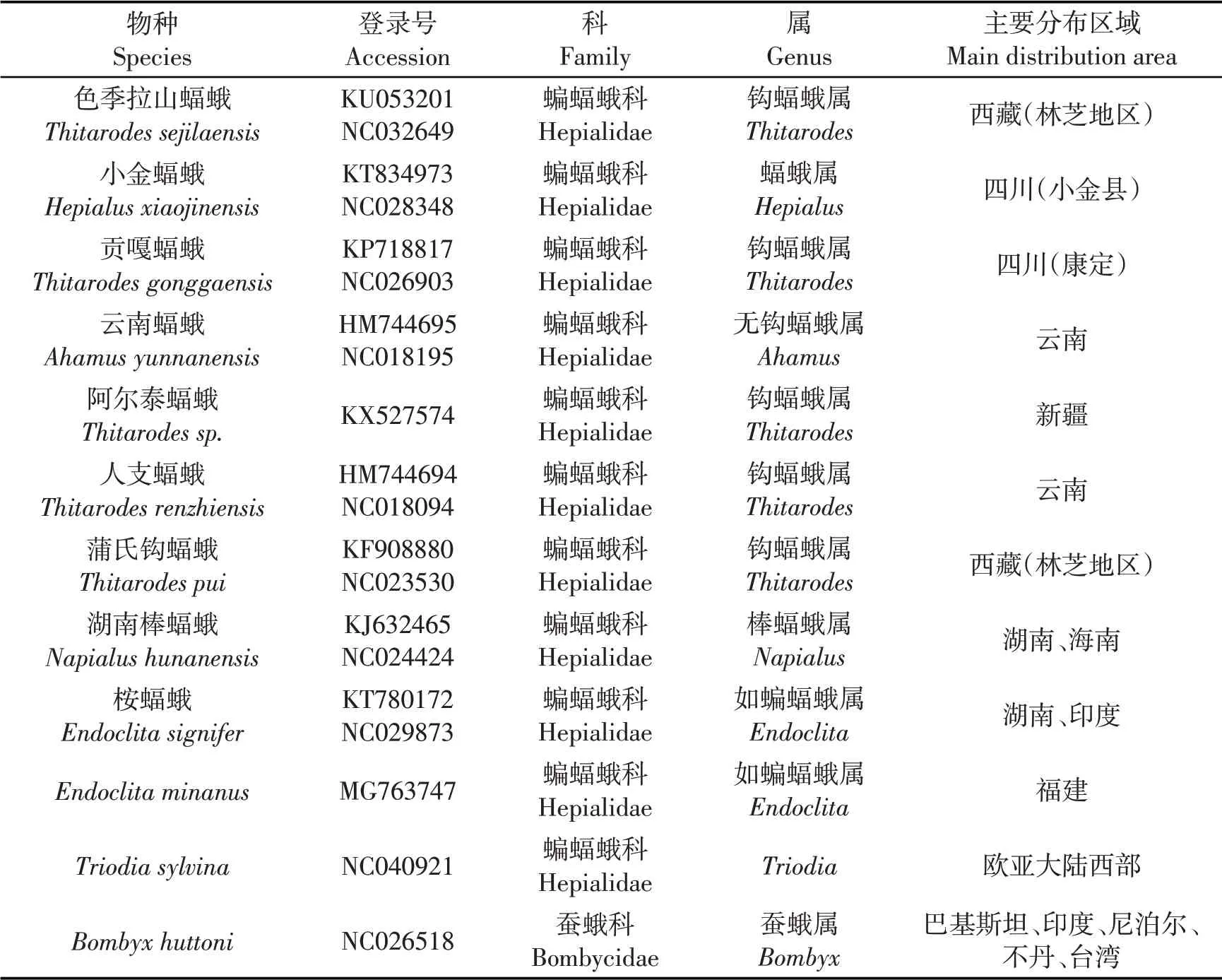
表1 本研究序列来源及信息

表1(续)
1.2 数据处理和系统进化分析
将从NCBI 中获取的序列保存为Fasta 格式,用ClustalX 2.1 软件进行序列对比. 然后将比对序列用MEGA 7.0软件统计不同蝠蛾科16S rRNA基因序列间的简约位点、可变位点、保守位点以及种间遗传间距,并构建系统进化树. 用DAMBE软件分析基因序列的碱基组成,并分析AT或GC偏斜性,计算公式为AT-skew=[A-T]/[A+T]和GC-skew=[G-C]/[G+C][39]. 本研究基于冬虫夏草、亚香棒虫草的寄主昆虫(蝠蛾类)及其他蛾类的16S rRNA基因用NJ法构建系统进化树. 为评估16S rRNA基因作为分类依据分子标记的可靠性,同时用相同物种线粒体基因组序列的16S rRNA 联合COⅠ基因构建ML 系统进化树. 将系统进化树中bootstrap自检值设置为1 000.
2 结果
2.1 序列碱基组成分析
将16S rRNA 基因序列进行对比排列后,长度为1 411 bp,其中保守位点1 054个,简约位点143个,可变位点310个. 通过计算可知,A、C、G、T碱基的平均含量分别为44.1%、5.3%、9.6%、40.0%. A+T的平均含量为85.0%,明显高于G+C 含量(15%). 由表2 可知,AT 和GC 的偏斜度均为正值且小于1,最小ATSkew值为0.005(T.sylvina),最大AT-Skew值为0.045(A.yunnanensis),最小GC-Skew值为0.26(T.sp.),最大GC-Skew值为0.326(H.xiaojinensis),说明11种蝠蛾碱基组成对AT显著偏移.

表2 11种蝠蛾的16S rRNA基因碱基组成
2.2 蝠蛾科11种蝠蛾的种间遗传距离
由表3可知,在11种蝠蛾中,种间遗传距离范围在0.005~0.117之间,人支蝠蛾(T.renzhiensis)与阿尔泰蝠蛾(T.sp.)和贡嘎蝠蛾(T.gonggaensis)的种间遗传距离为最小(0.005),T.sylvina与蒲氏钩蝠蛾(T.pui)间的遗传距离是最大(0.117). 虫草寄主蝠蛾间的遗传距离为0.005~0.51.T.sylvina与其他蝠蛾的遗传距离范围在0.099~0.117,相比其他蝠蛾间的遗传距离明显偏大.

表3 11种蝠蛾16S rRNA基因的遗传距离
2.3 系统进化分析
基于16S rRNA基因和COⅠ基因联合16S rRNA基因构建的系统进化树均明显分为两个大分支. 在NJ树(如图1)中第Ⅰ支由蝠蛾科昆虫包括虫草寄主蝠蛾及其他蝠蛾构成. 在虫草寄主蝠蛾中,人支蝠蛾(T.renzhiensis)与阿尔泰蝠蛾(T.sp.)聚为一支,与贡嘎蝠蛾(T.gonggaensis)及小金蝠蛾(H.xiaojinensis)聚成一簇,与色季拉山蝠蛾(T.sejilaensis)与蒲氏钩蝠蛾(T.pui)聚成的一支构成姊妹群. 云南蝠蛾(A.yunnanensis)位于此支系的基部(自展值100%). 7种虫草寄主蝠蛾构成一个独立的大支系(自展值为100%),表明属间亲缘关系较近. 如蝠蛾属的两种蝠蛾(Endoclita signifer和Endoclita minanus)单独聚类在一起,与湖南棒蝠蛾(N.hunanensis)和Triodia sylvina位于虫草寄主蝠蛾类群的基部. 而在ML树中(如图2),虫草寄主蝠蛾聚类与NJ树中一致,但非虫草寄主蝠蛾中N.hunanensis与T.sylvina单独形成一个支系,E.signifer和E.minanus聚类形成另一个单独支系. 两种树中第Ⅱ大支均由蚕蛾科、织蛾科、菜蛾科、卷蛾科、举肢蛾科和麦蛾科蛾类构成,NJ 树中蚕蛾科的家蚕Bombyx mori与Bombyx huttoni聚类形成一支,然后与菜蛾(Plutella xylostella)和卷蛾(Spilonota lechriaspis)形成一支. 举肢蛾科、织蛾科和麦蛾科聚类形成另一支.ML树中的第Ⅱ大支稍有不同,蚕蛾科单独聚类成一支,菜蛾(Plutella xylostella)和卷蛾(Spilonota lechriaspis)也聚类成一支,其余科的蛾类均单独形成一支(如图1和图2).
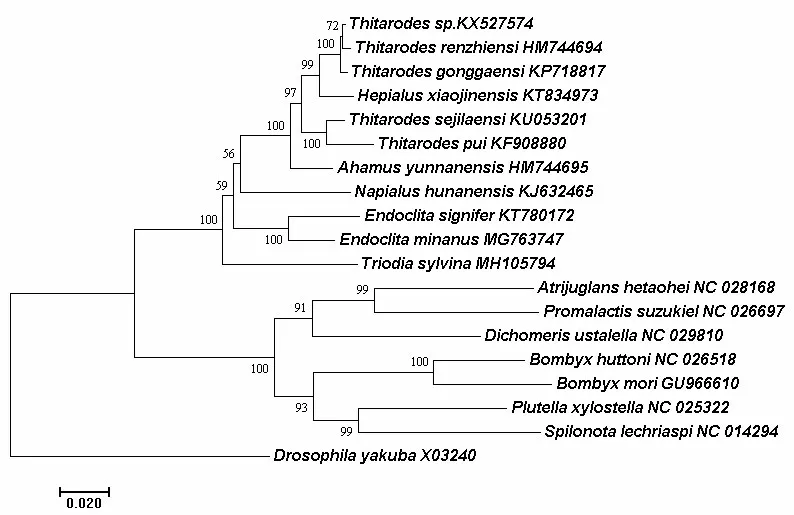
图1 基于线粒体16S rRNA基因构建的NJ树

图2 基于16S+COⅠ基因构建的ML树
3 讨论
16S rRNA基因之所以成为常用的线粒体分子标记,主要原因有如下几点:一是该基因存在于所有昆虫的体内;二是16S rRNA 基因的保守性和特异性比较高[37];三是该基因序列较长,约含有50 个功能域[37]. COⅠ基因在线粒体DNA中分布普遍,具有多态性较高、基因进化速率适中、适于通用引物扩增等特点,是昆虫分子系统学研究中较为理想的分子标记,该基因近年来在昆虫的群体遗传结构和系统进化方面有较多的研究[40]. 因此,在本研究中同时用虫草寄主蝠蛾16S rRNA基因联合COⅠ全基因构建系统进化树,以验证16S rRNA构建系统进化树的可靠性和有效性.
通过对蝠蛾科蝠蛾线粒体16S rRNA 基因序列进行分析,结果表明:序列中A+T 的平均含量为85.0%,G+C的平均含量为15.0%,碱基组成对A+T表现为显著的偏移. 种间的最小遗传距离为0.005(T.renzhiensis与T.sp.,T.renzhiensis与T.gonggaensis),说明三者的亲缘关系较其他蛾类更近. 将虫草寄主蝠蛾、其他蝠蛾及其他科蛾类的16S rRNA 基因全序列约1 400 bp个碱基作为系统进化树的分析位点. NJ系统树明显分为2大支(图1所示):第Ⅰ支由蝠蛾科昆虫包括虫草寄主蝠蛾及其他蝠蛾,7种虫草寄主蝠蛾最先聚为一支,其中人支蝠蛾(T.renzhiensis)、阿尔泰蝠蛾(T.sp.)与贡嘎蝠蛾(T.gonggaensis)同属钩蝠蛾属聚为一簇(种间遗传距离最小),小金蝠蛾(H.xiaojinensis)与3种钩蝠蛾属蝠蛾形成一个支系,说明小金蝠蛾(H.xiaojinensis)应为钩蝠蛾属,这一结果印证并支持Kang[11]等的研究结论. 这一支系与色季拉山蝠蛾(T.sejilaensis)和蒲氏钩蝠蛾(T.pui)形成的聚类簇互为姊妹群,而云南蝠蛾(A.yunnanensis)属无钩蝠蛾属,独立成一支. 以上结果与相关研究报道[11,18,29]所得结论一致. 同时发现,虫草寄主蝠蛾分布地域较近的也更趋于归为一类,如蒲氏钩蝠蛾(T.pui)和贡嘎蝠蛾(T.gonggaensis)均分布于西藏林芝地区. 湖南棒蝠蛾(N.hunanensis)是亚香棒虫草的寄主昆虫,并不是冬虫夏草的寄主昆虫,但两种虫草形态却极为相似,说明两种虫草寄主蝠蛾可能具有较近的亲缘关系. 进化树分析结果亦表明,在非虫草寄主蝠蛾中湖南棒蝠蛾(N.hunanensis)的确是与所有冬虫夏草寄主蝠蛾关系较近的一支,而其他蝠蛾与虫草寄主蝠蛾关系明显稍远一些. 本研究分析虫草寄主蝠蛾间的进化关系而得出的结论与Zou等[18]和Kang等[11]的观点基本一致. 本研究同时也用来源于相同物种线粒体基因组的COⅠ基因联合16S rRNA构建ML进化树(图2所示),与单独用16S rRNA基因构建的进化树结果相符,在一定程度上支持邹志文等[13]提出形态学分类的观点,即将蝠蛾属以雄性生殖器抱器瓣结构特征为依据分为4个属的合理性. 这也说明16S rRNA基因可以进行科属级分类,作为系统进化分析和分类的标记基因是有效且可信的.
本研究中样本数有所欠缺,分布区域不均,因此今后研究过程中为更全面具体地进行系统分类,还需要增加种群数量并结合其他分析技术手段,从而为虫草寄主昆虫的系统进化研究提供更全面的科学依据,同时为药用虫草资源的保护和利用奠定理论依据.
