水稻抗褐飞虱基因在物理图谱上的锚定
2020-03-20赵昶灵肖素勤程在全
顾 凡,陈 玲,陈 越,赵昶灵,肖素勤,程在全*
(1.云南农业大学 农学与生物技术学院,云南 昆明 650201;2.云南省农业科学院 生物技术与种质资源研究所,云南 昆明 650205)
褐飞虱(Nilaparvatalugens)是以水稻为食的单食性昆虫,从水稻中吸取汁液,当其为害严重时能引起稻飞虱群体大规模的暴发,给水稻种植区带来严重的破环,是亚洲最严重的水稻害虫之一。褐飞虱每年在中国的为害面积达40万hm2左右,为水稻种植总面积的50%左右[1]。水稻为禾本科稻属一年生栽培谷物,是我国重要的粮食作物,而在其种植中易发生褐飞虱危害,会导致水稻大片倒伏,从而形成枯孕穗或半枯穗,最终造成水稻大幅减产甚至颗粒无收。褐飞虱是典型的吸食维管束液的昆虫[2],其对中国和亚洲的稻米生产造成了很大的影响,特别是在易感水稻品种种植的地区,褐飞虱种群增长速度较快,对水稻生产构成了严重的威胁。选育和种植抗虫品种是褐飞虱综合防治的基础,抗褐飞虱品种的培育应用被认为是控制褐飞虱危害最经济和环境友好的途径[3]。自20世纪90年代以来,科学家们特别注意从野生稻中开发新的抗性基因,各国科学家历来都十分重视水稻抗褐飞虱资源的挖掘、研究和利用。近年来,利用分子标记技术也将一些抗褐飞虱基因进行了定位[4]。通过对与其紧密相连的分子标记的物理位置的查询,可以推断出分子标记在物理图上的大致位置。Bph14是从水稻中第一个被克隆的抗虫基因[5],从那时起,新基因的发现、鉴定、定位和克隆就被频繁报道。本研究将来自不同群体的褐飞虱抗性基因整合到水稻品种“日本晴”的测序图中,旨在为抗褐飞虱基因的克隆、分子标记选择和功能分析提供参考。
1 材料与方法
通过查阅大量文献(主要来源于http://www.Ricedata.cn/)以及在互联网上搜索,获得了褐飞虱抗性基因的鉴定、定位和克隆的总体情况,并获得了与褐飞虱抗性基因相关的分子标记。针对褐飞虱的定位基因使用不同的种群和标记。在构造的连锁图中,几乎没有发现其共同的标记,因此很难确定标记与基因之间的序列和距离。不过好在日本晴的测序工作已经完成,大部分的水稻标记都在其测序图上。因此,本文以日本晴的序列图作为参考图,整合不同来源的标记物和对褐飞虱的抗性基因。在整合过程中,在测序图上确定了与褐飞虱抗性基因相关的分子标记的序列和位置,并以该标记为参照标记了抗性基因在染色体上的物理位置。在本研究中,我们利用最新的连锁分子标记和对褐飞虱的抗性基因构建了一个新的褐飞虱抗性遗传图谱,该图谱是在www.gramene.org网站2009年发布的《Gramene注释的Nippon Bare Sequence》的序列图的基础上构建而成的。
2 结果与分析
2.1 抗褐飞虱基因鉴定、定位、克隆的总体情况
自20世纪70年代以来,随着分子标记技术的应用,人们对水稻抗褐飞虱基因的定位和克隆的研究越加频繁。到目前为止,水稻中有43个褐飞虱抗性基因,其中26个为显性基因,其余为隐性基因,这些基因均已得到国际注册和期刊报道。在这43个基因中,有38个抗性基因已完成定位工作,其分布在水稻7条染色体中,其中2号染色体1个,3号染色体5个,4号染色体15个,6号染色体6个,10号染色体1个,11号染色体1个,12号染色体9个。其中Bph3、Bph6、Bph14、Bph18、Bph26、bph29和Bph32等7个抗性基因已被克隆[6]。这38个抗褐飞虱基因大部分聚集在染色体的不同区域。
2.2 抗褐飞虱基因在染色体上的分布
经大量检索,将已鉴定并命名的水稻抗褐飞虱抗性基因绘制成表1,并通过染色体图谱的构建绘制成图1。从表1和图1可以看出,褐飞虱抗性基因Bph13(t)位于2号染色体上。位于3号染色体上也有一个褐飞虱抗性基因Bph13(t),但它们不是同一个基因。Bph13(t)这个基因的命名与Yang等[22]从药用野生稻中鉴定出的基因是重复的,因此,为了区分彼此,本文将2号染色体上的基因命名为Bph13(t)-1,将3号染色体上的基因命名为Bph13(t)-2。此外,在第3号染色体和第12号染色体上均发现了抗性基因bph19(t),但两者也并非同一个基因,为区分彼此,本文特将其分别命名为bph19(t)-1、bph19(t)-2。基因的重复命名也同样发生在位于第3号和第4号染色体上的Bph31(t),经研究两者也并非同一基因,故分别将其命名为Bph31(t)-1、Bph31(t)-2。位于第6号染色体上的基因bph20(t)与bph25(t),由于前期发现人及其所用研究方法的不同,得以分别命名,但经后期研究发现两者是同一个基因。同样先后发现的位于第10号染色体的基因bph21(t)、bph26(t),现在研究结果表明两者为同一基因。
褐飞虱抗性基因Bph13(t)-1位于2号染色体上。刘国庆等[7]利用RFLP和SSR等分子标记将Bph13(t)-1定位在2号染色体上,位于两个微卫星标记RM240和RM250之间。它在2号染色体上的物理距离是31.50~32.78 Mbp。该基因的发现和定位将有助于对水稻褐飞虱抗性的改良[7]。
在3号染色体上鉴定出5个抗褐飞虱基因,分别为bph11、Bph13(t)-2、Bph14、bph19(t)-1和Bph31(t)-1。Bph13(t)-2[22]来源于紧穗野生稻中,位于3号染色体上,定位于RG100和RG191标记之间。Bph14[2]来源于药用野生稻,其与Feb-76标记共分离;该基因cDNA全长为9576 bp,包含3个外显子,编码由1323个氨基酸组成的蛋白质产物,产物包括螺旋结构域、核苷酸结合域和富含亮氨酸的重复序列(CC-NB-LRR)。重组自交系RI35与台中1号中的Bph14基因序列差异很大,其中CC域和NB域较为保守,而LRR具有独特的差异[8]。bph19(t)-1来源于水稻品种AS20-1,被定位在RM517与RM218标记之间,其位于第3号染色体的物理距离在6.16~8.40 Mbp[27]。Bph31(t)-1来源于品种CR2711-76,对褐飞虱生物型Ⅳ有很好的抗性,其被定位于RM251和RM2334之间[41]。
在第4号染色体上定位了15个抗褐飞虱基因,即Bph6、Bph12(t)、bph12、Bph15、Bph17、bph18(t)、Bph20(t)、bph22(t)、bph23(t)、Bph24(t)、Bph27(t)、Bph30、Bph31(t)-2、Bph33和Bph34。Bph6位于4号染色体Y37标记处,约21.4 Mbp的位置[16]。来源于阔叶野生稻的显性基因Bph12(t),被定位于水稻第4号染色体上的RM261和RM8213之间,且位于第4号染色体上4.44~6.57 Mbp之间[22]。bph12位于4号染色体上的G271和R93之间,在染色体上的物理距离为20.34~21.31 Mbp[21]。宋丁丁等[23]将Bph15定位于第4号染色体的分子标记M1(STS)位置,其位于第4号染色体上物理距离6.94 Mbp处。Sun L H等[24]通过连锁作图和QTL分析将Bph17定位到4号染色体短臂上的RM8213和RM5953标记之间,其在染色体上的物理距离为4.45~9.38 Mbp。bph18(t)位于RM6506和RM273标记之间,在染色体上的物理距离为24.05~25.05 Mbp[26]。Rahman等[29]将Bph20(t)定位于水稻第4号染色体短臂区,它与标记B43共分离,位于染色体物理距离8.76 Mbp处。bph22(t)被定位于第4号染色体RM8212和RM261标记之间[8]。来源于广西野生稻的Bph24(t)位于两个标记YZ16611和YZ26之间[30]。Huang等[36]从普通野生稻中鉴定出Bph27(t),并将其定位在第4染色体上的RM16852标记上,其大概位于染色体物理距离19.18 Mbp的位置。目前众多抗褐飞虱基因分布于4号染色体上。众所周知,抗病基因也集中在相同染色体的区域[9]。
在6号染色体上鉴定出6个抗褐飞虱基因,分别为Bph3、bph4、bph20(t)/bph25(t)、Bph25、bph29和Bph32。Bph3[13]和bph4[14]位于水稻第6号染色体短臂的同一区域,这2个基因可能紧密相连或是等位基因。从广西野生稻上挖掘的bph20(t)/bph25(t)基因位于BYL7与BYL8标记之间,在染色体上物理距离0.47~0.53 Mbp[26]。Myint K K M等[34]在水稻品种ADR52中发现的抗褐飞虱Bph25基因被定位在第6号染色体S00310标记上,该标记是Bph25基因的共分离标记,位于第6号染色体物理距离0.21 Mbp处。bph29位于标记RM540和RM435之间,在第6号染色体上的物理距离为0.38~0.53 Mbp[38]。另外,据文献报道,抗褐飞虱bph29基因含有B3结合域,能激活水杨酸信号通路,抑制茉莉酸/乙烯通路。
褐飞虱抗性基因bph21(t)/bph26(t)位于第10号染色体上。bph21(t)/bph26(t)来源于广西野生稻,位于标记RM222和RM244之间,其在第10号染色体上的物理距离为2.62~5.00 Mbp[30]。
褐飞虱抗性基因Bph28位于第11号染色体上。Bph28来源于水稻品种DV85,该基因被定位在标记Xue17处,该标记与基因Bph28共分离,位于染色体物理距离9.35 Mbp位置[37]。
在第12号染色体上定位了9个抗褐飞虱基因,即Bph1、bph2、bph7、Bph9、Bph10、Bph18(t)、bph19(t)-2、Bph21(t)和Bph26。第一个被鉴定出的抗性基因Bph1位于12号染色体的pBPH9标记处,在染色体物理距离22.86 Mbp位置[11]。bph2被定位于第12号染色体上SSR标记RM7102和RM463之间[12]。bph7位于12号染色体上RM3448和RM313之间,在染色体上的物理距离为19.95~20.87 Mbp[17]。苏昌潮等[19]对Bph9进行了连锁分析,发现Bph9基因位于水稻12号染色体的InD2和RM28466标记之间,其在染色体上的物理距离为22.85~22.97 Mbp。抗褐飞虱Bph9基因在表达起作用时会编码一种罕见核苷酸,引起细胞死亡,在水稻植株中激活水杨酸和茉莉酸信号通路,从而对褐飞虱产生抗性,抑制褐飞虱的取食[10]。Bph10被定位于第12号染色体上标记RM260和RM17之间[20]。抗性基因Bph18(t)来源于澳大利亚野生稻,位于12号染色体长臂末端的标记KC1/7312和T4A(KT)之间,其位于染色体物理距离22.88 Mbp处[25]。Bph21(t)基因位于12号染色体长臂端194.0 kb的区间内,介于两个标记RM3726和RM5479之间,在染色体上的物理距离为23.27~24.41 Mbp[29]。Bph26位于12号染色体上,其与RM5479标记共分离,在染色体上的物理距离约为24.41 Mbp[35]。
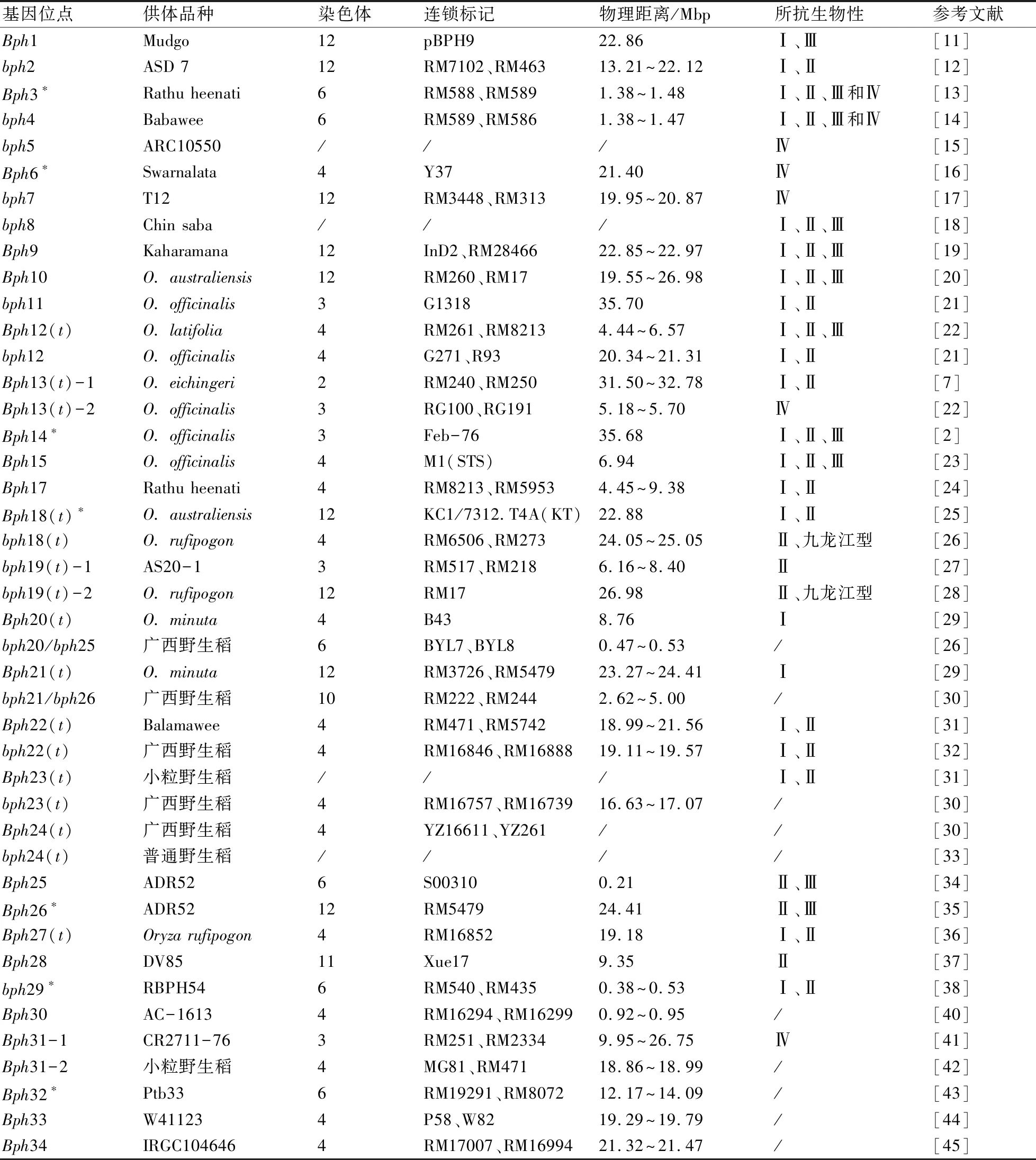
表1 已鉴定并命名的水稻抗褐飞虱抗性基因
注:“*”表示该基因已被克隆。
3 讨论
3.1 抗褐飞虱基因命名混乱的问题
到目前为止,已有至少43个水稻褐飞虱基因被报道,其中38个基因已被定位,其中一些基因是紧密相连的或是等位基因。前期由于供者亲本和标记方法的不同,在不同的研究机构中鉴定出的同一基因可能存在一定的差异。例如位于水稻6号染色体上的bph20(t)与bph25(t)基因就是由于前期发现人和供体材料的不同,得以分别命名,后经研究发现两者是同一基因。位于10号染色体上的bph21(t)与bph26(t)基因,亦是这种情况。再者不同基因的命名相同现象也时常发生,比如位于2号和3号染色体上均有基因Bph13(t),但两者并非同一基因,为了区分彼此,本文将其分别命名为Bph13(t)-1和Bph13(t)-2;在3号、12号染色体上的bph19(t)基因与3号、4号染色体上的bph31(t)基因也是这种情况。抗褐飞虱基因命名混乱的问题给科研工作带来了不良的影响,沟通交流时容易造成歧义,给学术交流制造障碍,也会造成财力、物力上的浪费。因此在今后抗褐飞虱基因的命名上应努力做到基因命名的标准化,实现命名的统一,以便在今后的科研工作中能够更加快捷、有效的沟通,也有助于其他领域的科学家联合开展工作,共享研究成果。
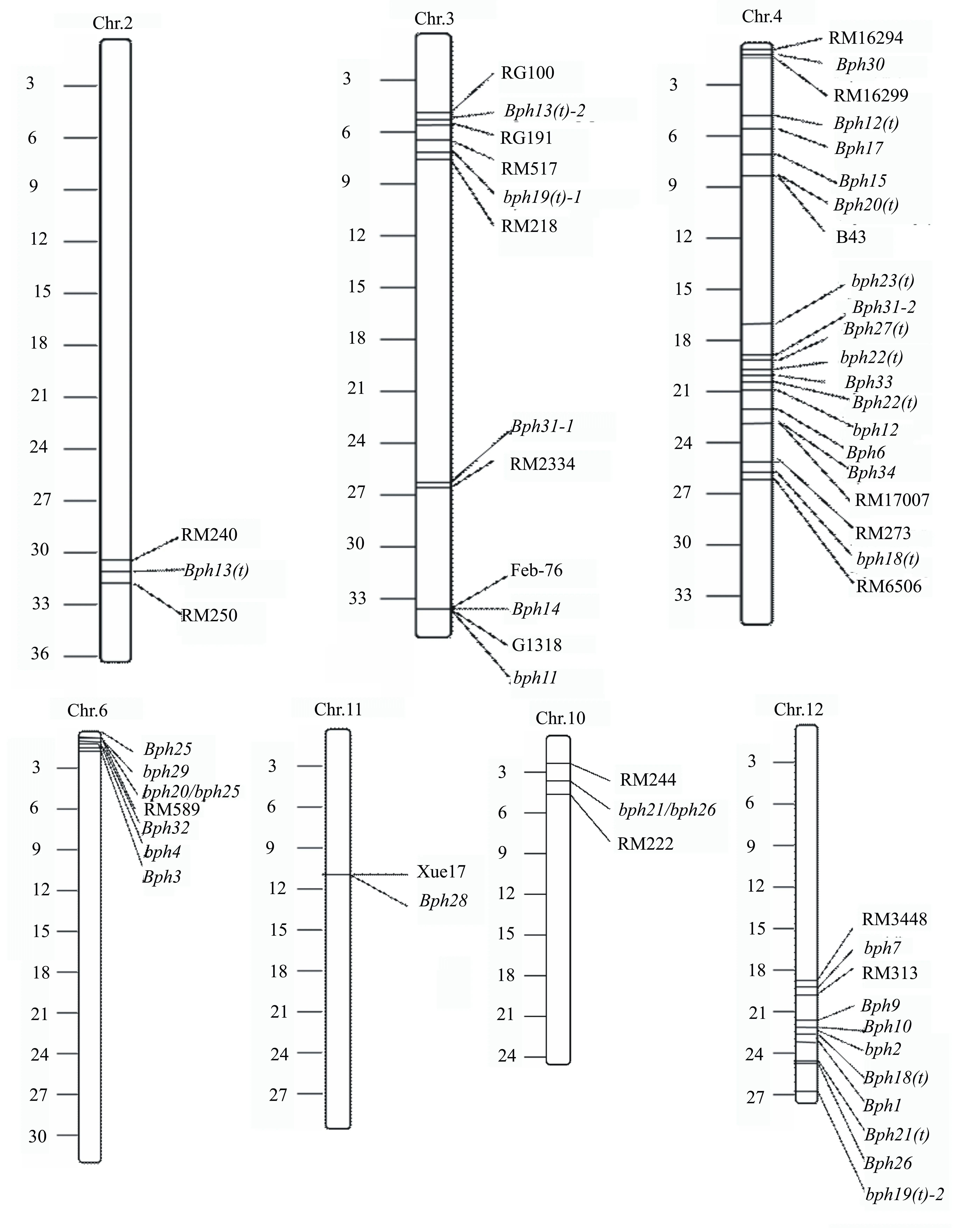
左边刻度为3×106个碱基对;右侧文字为标记和基因的相对位置。图1 水稻抗褐飞虱基因在物理图谱上的位置
3.2 抗褐飞虱基因成簇分布于染色体上
据前人研究报道,有些抗褐飞虱基因并不是随机、无序地分布于水稻各个染色体上,这38个褐飞虱抗性基因大部分以基因簇的形式分布在水稻的不同染色体区,其中在水稻4、6、12号染色体上形成了较大的基因簇。这些抗褐飞虱基因紧密连锁,成簇状集中排列在同一条染色体的某一特定区域。这些褐飞虱抗性基因彼此关联,相互配合,共同发挥作用。相比于褐飞虱单个抗性基因,抗褐飞虱基因簇的存在使得其基因间配合更加密切、高效,能更好地发挥其生物功能。除了这3个比较大的基因簇外,在其他染色体上也分布着比较小的基因簇。在38个已鉴定和定位的褐飞虱抗性基因中,有大量基因位于特定的基因簇中。但是这些基因簇的分布也增加了等位基因测定的难度和基因间抗性谱的差异,因此很难判断抗褐飞虱基因簇中各个基因彼此间的差异情况。在未来,基因精细定位技术的不断发展、基因克隆和基因间抗性谱测定的精准性提高,都将帮助我们探讨抗褐飞虱基因簇中各个基因的结构以及彼此之间的关系,明确各个抗褐飞虱基因的生物功能。
3.3 抗褐飞虱基因在育种上的应用
在常规育种中,通过有性杂交培育褐飞虱抗性品种是目前最常见也是应用最广的方法。目前许多褐飞虱抗性品种已被培育和推广,但是在常规育种中,还存在许多问题:一方面耗时较长,在短时间内无法获得好的抗性品种;另一方面随着时间的推移,一些高抗水稻品种会出现抗性退化的现象,并且可能出现褐飞虱新的“生物型”,导致辛苦培育的品种失去抗性。因此,如何培育具有持久抗性的抗褐飞虱水稻品种逐渐成为当前科学家和技术人员面临的难题。
在分子标记辅助(marker-assisted selection)以及基因工程育种方面,许多抗褐飞虱基因抗谱窄,并且具有连锁累赘和隐性特征,使得基因的转育效率极低[46]。目前除了基因Bph1、bph2、Bph3、Bph14和Bph15外,尚未能有效利用其他的抗褐飞虱基因[47]。有效利用已知抗性基因,继续深入挖掘更多抗性基因,是今后分子标记育种工作的重要内容。前人的研究结果表明,在利用分子标记辅助选择抗褐飞虱品种时,含单个抗性基因的品种的抗性没有含两个或多个抗性基因的品种强,这就提醒我们今后利用分子标记辅助选择培育抗性品种时,要有目的地聚集抗虫基因和QTL,寻找与抗虫基因紧密相连或分离的分子标记,尽可能多地培育包含两个或两个以上的抗性基因品种,增加品种的耐抗性,延缓抗虫品种的退化时间,防止出现新的褐飞虱生物型[48]。当前许多优良的褐飞虱抗性基因是在野生稻种上被发现挖掘的,因此,育种工作者必须充分利用野生水稻尤其是药用野生稻,以及含有褐飞虱抗性基因的突变体植株,以此来加强对褐飞虱新抗性基因的开发挖掘。目前水稻褐飞虱的大部分抗性基因都已被定位,这大大提高了褐飞虱基因转移的效率,有助于褐飞虱抗性基因的利用[49]。总的来说,利用分子标记对水稻育种材料进行筛选培育,能够准确判断其是否带有抗褐飞虱基因,以及所带抗性基因的类型。目前,我国已成功利用分子标记辅助选育出一批抗褐飞虱的水稻新品种,褐飞虱对水稻的危害得到了较大的控制。
如何将水稻常规育种方法和分子标记辅助选择方法相互结合,使两者取长补短,既能缩短育种周期,又能准确高效地筛选培育出高抗褐飞虱的品种,这是育种家们要面对的难题,也是今后研究工作的努力方向。
本研究将38个已定位的抗褐飞虱基因整合到日本晴的序列图中,在其物理图谱上可以找到基因的连锁标记及其区域;通过检索GRAMENE、RGP网站可以获得连锁标记之间的片段。水稻抗褐飞虱基因在物理图谱上的锚定,为今后基因的精细定位、基于图位克隆和通过分子标记辅助选择培育新品种打下了基础,有助于水稻抗褐飞虱基因的系统研究。
