夏休闲期复种饲料油菜提升后作冬小麦产量品质、土壤肥力及周年经济效益研究
2020-03-17李军辉杨珍平高志强
李军辉 王 凯 杨珍平 高志强
(山西农业大学农学院,山西 太谷 030801)
山西省位于黄土高原东缘,属大陆性半干旱气候,干旱、土壤肥力低下是限制全省小麦生产的重要因素之一[1-3]。晋中地区是山西省小麦生产的重要区域,仅次于南部临汾、运城主产区。从小麦生态气候区划来看,晋中麦区属北方晚熟冬麦区[4],从小麦品质区划来看,则隶属强中筋优质麦区。该区年平均气温6.6~10.8℃,年积温3 350.1℃,年日照时数2 330.8~2 704.3 h,种植制度多为一年一作。每年9月25日秋分前后种植冬小麦,翌年6月20-25日收获,整个生育期260 d 左右,7-9月为休闲期。该区全年平均降雨量约458 mm,而休闲期降水约占全年降水的60%以上。因此,前人对休闲期覆盖保水方面进行了大量研究。贺立恒等[5]和孙敏等[6]研究表明,在休闲期进行土壤深松有利于提高播前土壤含水量,有利于小麦生长;任爱霞等[7]研究表明,夏休闲期覆盖地膜同样能有效提高土壤蓄水保墒能力,提高小麦产量和籽粒品质;此外,提前深施有机肥,深松配套膜际条播方式等均可以明显提高土壤蓄水保水以及小麦产量[8-10]。总之,旱地小麦夏休闲期三提前(提前土壤耕作、提前施用有机肥、提前秸秆或地膜覆盖)技术,是提高旱地小麦生育期间0~300 cm 土层土壤含水量、提高籽粒水分高效利用的重要措施。在夏休闲期无论是采用不同耕作措施,还是覆盖地膜,都仅提高了土壤水分,而光热资源没有得到充分利用。根据姚致远等[11]在陕西长武的研究,夏休闲期种植油菜绿肥不仅起到了对土壤水分的保护作用,而且充分利用了夏休闲期的光热资源,提高了光能利用率和土地利用率。本研究拟将饲料油菜绿肥引入晋中麦区,探讨其对后作小麦生长及经济效益的影响,旨在为合理利用休闲期、提高晋中麦区经济效益提供理论依据。
1 材料与方法
1.1 试验地概况
本试验于2015年6月-2016年6月在山西省晋中市太谷县山西农业大学实验田内进行。试验田土壤类型为褐土,土质中壤土,0~40 cm 耕层土壤养分含量为有机质13.25 g·kg-1、全氮1.89 g·kg-1、全磷198.5 mg·kg-1、速效氮57.5 mg·kg-1、速效磷11 mg·kg-1、速效钾135 mg·kg-1,pH值8.0。
1.2 供试材料
饲料油菜品种:华油杂62号,甘蓝型半冬性波里马细胞质雄性不育系杂交种,春油菜晚熟组全生育期140 d 左右,由华中农业大学、湖北国科高新技术有限公司提供。
冬小麦品种:CA0547,冬性,强筋,由中国农业科学院作物科学研究所育成。
1.3 试验设计
2015年6月28日,前茬小麦收获后,撒播饲料油菜,播种量22.5 kg·hm-2,播种面积300 m2,设不种油菜为空白对照(CK0)。重复3 次。9月14日,在上述每个重复的饲料油菜田中取3个0.667 m2的样段,将地上部植株收获,测定鲜重和干重(经测定平均鲜重4 443.3 kg·666.7 m-2,平均干重543.3 kg·666.7 m-2),将植株晾干粉碎,测定植株N、P含量(经测定平均N含量为24.05 kg·666.7 m-2,平均P含量为2.38 kg·666.7 m-2)。之后,将每个重复的饲料油菜田平均划分成2个小区,其中一个小区将油菜植株的地上部分全部收获(CK1),另一个小区将油菜植株的地上部分全部粉碎还田(T)。9月26日,将上述所有地块统一进行旋耕处理,并机械条播后作小麦,行距20 cm,播种量15 kg·666.7 m-2。
1.4 测定指标与方法
1.4.1 植株农艺性状及籽粒蛋白质组分含量测定 于后作小麦三叶期(2015年10月16日)、越冬期(2015年12月1日)、返青期(2016年3月8日)、起身期(2016年4月2日)、拔节初期(2016年4月15日)、拔节中期(2016年4月23日)、孕穗期(2016年5月1日)、抽穗期(2016年5月4日)、扬花期(2016年5月11日)进行随机取样,每个处理取10株,测其株高、绿叶数、旗叶长宽、倒二叶长宽以及整株干重和穗干重。再于开花期选择同天开花、大小均匀的穗挂牌标记,于花后5、10、15、20、25、30 d 取样,每次取30个穗和10 片旗叶。取部分穗于105℃杀青30 min,80℃烘干至恒重,然后粉碎,最后采用连续提取法测定籽粒清蛋白、球蛋白、醇溶蛋白和谷蛋白含量。采用半微量凯氏定氮法测定含氮量,再乘以5.7 即为蛋白质含量[12]。每个处理再取10个茎,测其旗叶长宽、倒二叶长宽、单茎干重和穗干重。
1.4.2 产量测定 在小麦成熟期,每个处理随机取3个面积为0.667 m2的小区进行单独收获,并计算每个小区的总穗数、穗粒数、千粒重和实际产量。
1.4.3 植株N、P含量测定 采用植株NPK 联合测定法[13]。将三叶期、越冬期、返青期、起身期、拔节初期、拔节中期、孕穗期、抽穗期、扬花期以及花后5、10、15、20、25、30 d 所取单株或单茎样品,烘干,粉碎,称重,经H2SO4-H2O2消煮,全氮测定采用半微量凯氏定氮法,全磷测定采用钒钼黄比色法。
1.4.4 土壤酶活性、土壤有机质含量及土壤相对含水量的测定 分别于上述后作小麦的拔节初期(2016年4月15日)和孕穗期(2016年5月1日),在每个处理随机选3个样点,在小麦植株根轴部位,用直径8 cm的螺旋式土钻,按0~10、10~20、20~40、40~60、60~80和80~100 cm的垂直土层依次分层获取土样,所取土样分成2份,1份装入铝盒用于土壤相对含水量的测定,1份装入自封塑料袋以备土壤酶活性和土壤有机质含量的测定。
采用烘干直接称重法测定土壤相对含水量。
将上述土样风干,粉碎,按土壤养分测定的要求过0.25 mm 筛,采用重铬酸钾容量法——外加热法[12]测定有机质含量;按土壤酶活性测定的要求过1 mm 筛,采用靛酚比色法测定土壤脲酶活性[14],酶活性以培养24 h后每克干土中NH3-N的质量变化表示;采用3,5-二硝基水杨酸比色法测定土壤蔗糖酶活性[12],酶活性以培养24 h后每克干土中葡萄糖的质量变化表示。
1.5 周年经济效益计算
根据当年饲料油菜和小麦的实际产量与收购价格,计算产出值;根据当年农机作业、种子、肥料、农药、人工等投入,计算生产成本费用;从产出值中扣除生产成本后,即为冬小麦-油菜的周年经济效益。
公式计算如下:

1.6 数据统计与分析
采用Microsoft Office Excel 2007 制表作图,并用SAS 9.0软件对数据进行ANOVA 方差分析和多重比较。
2 结果与分析
2.1 麦后复种饲料油菜对后作冬小麦生长发育的影响
2.1.1 麦后复种饲料油菜对后作冬小麦抽穗前单株绿叶数及株高的影响 由图1可知,从三叶期到孕穗期,随着生育期的延长,后作冬小麦单株绿叶数及株高均呈逐渐增加的趋势;就单株绿叶数而言,从三叶期至越冬期为第一次增长高峰期(P<0.05),从起身期至拔节初期为第二次增长高峰期(P<0.05),从拔节初期开始各处理之间的差异增大,以T的单株绿叶数最多,且显著高于其他处理(P<0.05)。而株高的显著增加发生在起身期至孕穗期,尤其起身期至拔节初期增长幅度更大,但3个处理间仅在拔节初期达到差异显著水平,且T的株高在整个拔节期均显著低于其他处理(P<0.05),其原因可能是还田处理需要消耗部分土壤营养用于油菜腐解,同时还田处理促进了单株分蘖增多,植株生长健壮,因而株高偏低。

图1 不同处理下小麦抽穗前单株绿叶数(A)和株高(B)的变化曲线Fig.1 Changing curves of green leaves (A) and plant height (B) per wheat plant in the different treatments
2.1.2 麦后复种饲料油菜对后作冬小麦功能叶叶面积的影响 本研究中,扬花期前调查的是单株功能叶片(旗叶和倒二叶,表1)的长、宽和面积;花后灌浆期调查的是单茎功能叶片(旗叶和倒二叶,表2)的长、宽和面积。
由表1可知,花前单株功能叶片的长、宽和面积受生育时期的影响(旗叶的变异系数约33.34%~62.73%,倒二叶的变异系数约15.65%~42.91%)明显大于受处理的影响(旗叶的变异系数约1.45%~27.81%,倒二叶的变异系数约0.33%~10.36%),与上述单株绿叶数和株高的情况类似。不同处理对功能叶片叶宽的影响(旗叶的变异系数为1.53%~20.66%,倒二叶的变异系数为2.11%~10.36%)明显大于对叶长的影响(旗叶的变异系数为1.45%~4.26%,倒二叶的变异系数为0.33%~3.80%)。整体而言,旗叶的叶面积在扬花期达到最大值(CK0叶面积除外),孕穗后变化不大;尤其叶宽和叶面积,处理间的差异主要表现在起身期-拔节中期,其次是扬花期,在孕穗期、抽穗期3个处理间无显著差异(P>0.05)。倒二叶的叶长、叶宽及叶面积增长主要在起身期-拔节初期,各处理间的差异大多不显著(P>0.05)。
由表2可知,随着灌浆期的推进,小麦功能叶的叶长、叶宽及叶面积呈缓慢降低的趋势。总体而言,种植油菜处理能在一定程度上保持灌浆期旗叶叶面积,对倒二叶叶面积的影响也不大。
2.2 麦后复种饲料油菜对后作冬小麦产量及产量结构的影响
由表3可知,3个处理的产量结构差异主要表现在穗粒数上,为T>CK0>CK1(P<0.05);总穗数和千粒重差异不显著(P>0.05)。从实际产量来看,T 较CK0增产5.7%,但两处理差异不显著(P>0.05),CK1产量较CK0显著降低。
2.3 麦后复种饲料油菜对后作冬小麦植株氮磷、干物质积累及籽粒蛋白质含量的影响
2.3.1 对植株氮磷积累的影响 由表4可知,生育时期对小麦植株N、P含量积累的影响远大于不同处理的影响。花前小麦植株N、P含量呈先升高后降低的变化趋势,其中N含量在孕穗期达到最大值(P<0.05),P含量在返青期达到最大值。处理之间比较,返青期之前以CK0的植株N含量最高(P<0.05),起身期至拔节中期以T的植株N含量最高(P<0.05),可能原因是冬前还田处理需要消耗一部分营养用于油菜腐解,而在返青期至拔节期的分蘖大量发生期,T 因油菜腐解后增加了土壤无机N含量,从而促进了根系吸收及茎叶N 积累。而对于植株积累P含量,T 则不具备优势。

表1 不同处理下小麦返青期至扬花期单株功能叶面积的变化Table1 Change of functional green leaf area per wheat plant from reviving stage to flowering stage of the different treatments

表2 不同处理小麦灌浆期单茎功能叶面积的变化Table2 Change of functional green leaf area per wheat stem in the different treatments
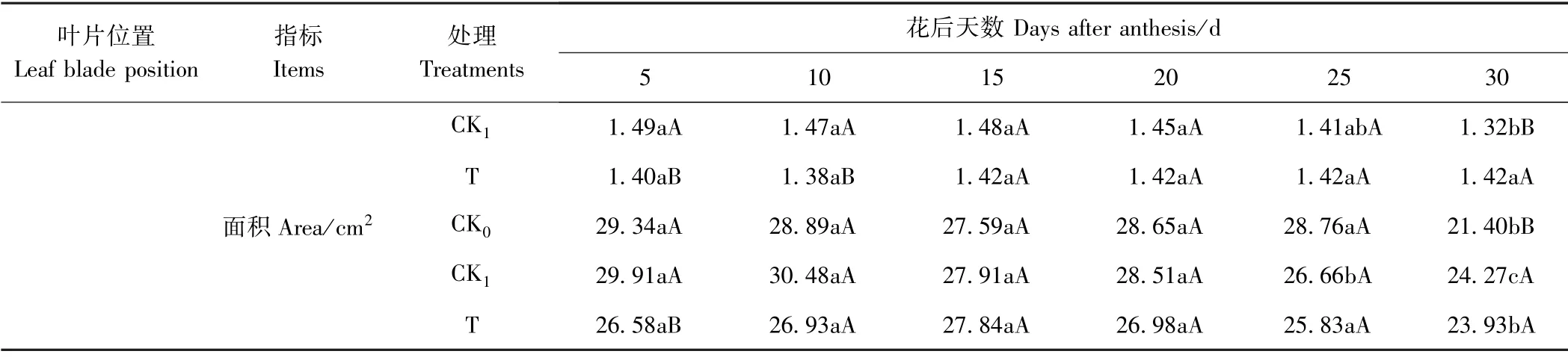
表2(续)

表3 不同处理对小麦产量及产量结构的影响Table3 Effects of different treatments on wheat yield and yield composition
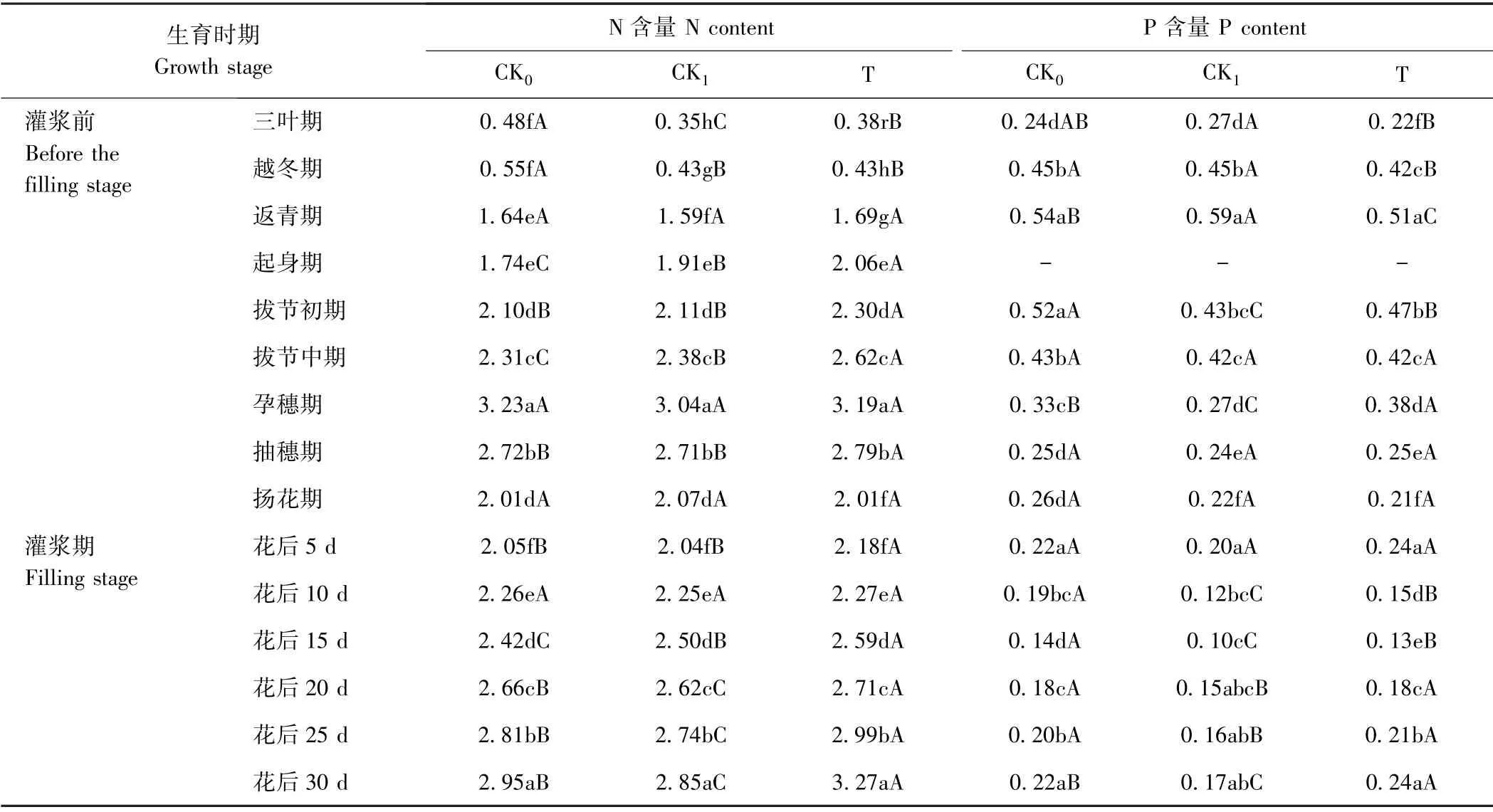
表4 不同处理对小麦植株氮磷含量的影响Table4 Nitrogen and phosphorus contents of wheat plant /(g·kg-1)
观察灌浆期的单茎N、P含量,发现随着灌浆期的推进,单茎N 积累呈逐渐升高趋势,花后30 d 达到最大值(P<0.05);单茎P 积累呈先降低后升高的趋势,花后15 d 最低,花后5 d和30 d 差异不显著(P>0.05)。处理之间比较,T的单茎N 积累较高,其单茎P 积累在花后20~30 d 也最高(表4)。总之,麦后复种饲料油菜有利于后作小麦植株或单茎N、P 积累量的提高。
2.3.2 麦后复种饲料油菜对后作冬小麦灌浆期干物质积累影响 在灌浆期,随着花后同化产物积累及植株干物质向穗部转移,穗部干重呈逐渐增加趋势(图2-B),而植株干重呈下降趋势(图2-A)。本试验中,花后5~20 d是植株干物质快速运转期,花后20~30 d是穗部干物质快速增长期,说明花后20~30 d 穗部干物质积累主要源于光合产物积累。处理之间比较,T的植株干物质运转及花后光合产物积累均明显高于其他两个处理。

图2 不同处理下小麦灌浆期植株干重(A)、穗干重(B)变化趋势Fig.2 Line diagrams of dry weights per plant and per panicle for wheat under the different treatments in the filling period
2.3.3 油菜还田对小麦灌浆期籽粒蛋白质积累特性的影响 由表5可知,随着灌浆期的推进,小麦籽粒蛋白质总含量整体呈现先下降后上升的趋势,在花后10 d 到达最低点,之后持续升高,在花后30 d 达到最大值,其中T的蛋白质含量显著高于CK0和CK1(P<0.05)。观察各蛋白组分含量随灌浆进程的变化趋势可知,清蛋白含量呈持续下降趋势;球蛋白含量与蛋白质总含量趋势类似,呈先下降后上升的趋势,在花后15 d 达到最低;而醇溶蛋白和谷蛋白含量从花后5 d开始持续增长,在花后30 d 达到最高。总体来说,在灌浆初期(花后0~10 d),小麦籽粒中以清蛋白和球蛋白为主要成分,进入灌浆高峰期(花后10~20 d)后,清蛋白含量降低,球蛋白含量先降低后升高,醇溶蛋白和谷蛋白含量增加。这种变化可能与小麦品种有很大关系,CA0547是一种强筋小麦,湿面筋含量较高(32%~35%),而醇溶蛋白和谷蛋白水合后形成面筋,是决定小麦面筋含量的重要因素,因此,在灌浆后期谷蛋白和醇溶蛋白积累增多。处理间比较,T 明显提高了灌浆后期(花后20~30 d)籽粒醇溶蛋白和谷蛋白含量,提升了小麦籽粒品质。
2.4 麦后复种饲料油菜对后作冬小麦土壤有机质、酶活性及水分状况的影响
本试验选择后作冬小麦分蘖发生高峰期(拔节期,2016年4月15日取样)和分蘖两级分化期(孕穗期,2016年5月1日取样)进行后作小麦根际土壤有机质含量(图3-A)、脲酶活性(图3-B)、蔗糖酶活性(图3-C)以及土壤含水量(图3-D)的测定。对上述各指标分别作生育时期(a)、还田处理(b)及土层深度(c)的三因素方差分析及多重比较(表略),结果表明,试验模型总方差均达到差异极显著水平(P<0.001 或P<0.000 1,R2=0.877 1~0.962 5),分析可靠。
由图3-A可知,拔节期的有机质含量明显高于孕穗期,处理之间以T的有机质含量最高,说明油菜秸秆还田腐解后,可能为土壤带来了大量腐殖质;土层之间以0~10 cm 土层的有机质含量最高,0~40 cm 土层平均有机质含量达到10 g·kg-1以上;在拔节期和孕穗期,随着土层加深,3个处理的有机质含量均逐渐递减。此外,在麦收后复种油菜前,0~20、20~40、40~60、60~80、80~100 cm 土层的播前土壤有机质含量分别为11.25、10.01、8.42、6.73、6.16 g·kg-1,表现为随着土层加深有机质含量降低。通过比较播前土壤有机质含量和后作小麦拔节期、孕穗期土壤有机质含量,发现,T 在0~40 cm 土层的有机质含量均高于播前土壤;CK0除0~20 cm 土层的有机质含量高于播前土壤外,其余土层均低于播前土壤;CK1则在拔节期0~40 cm土层的有机质含量高于播前土壤,孕穗期0~20 cm土层的有机质含量高于播前土壤。可见油菜还田对于土壤有机质含量有一定的提升作用。

表5 小麦灌浆期籽粒蛋白质及蛋白质组分含量Table5 Grain protein and protein component contents of winter wheat /%
由图3-B可知,孕穗期的脲酶活性明显高于拔节期;在拔节期和孕穗期,随着土层加深,3个处理的后作小麦根际土壤脲酶活性均以10~20 cm 土层最高(P<0.05)(CK0和CK1在孕穗期未达到显著),随土层加深,脲酶活性逐渐降低;0~10 cm 表层土壤的脲酶活性明显低于10~20 cm 土层,而CK1在0~10 cm 表层土壤的脲酶活性甚至近于或低于40~60 cm 土层。处理之间,T的平均脲酶活性明显高于CK0和CK1;T 在0~60 cm 土层的根际土壤脲酶活性最高,表明油菜还田可以明显提高该层次土壤中脲酶活性,从而有利于吸收土壤中的N 素;T 在60 cm 以下土层的脲酶活性不及CK0或CK1,可能是由于油菜还田深度在0~40 cm土层范围内,促进了该层及其邻近土层40~60 cm 土层的微生物繁衍、脲酶活性提高,而60 cm 以下土层反而有所降低,这有待进一步深入研究。CK1在拔节期的脲酶活性明显低于CK0,而在孕穗期其在10~60 cm土层的脲酶活性则高于CK0,可能是由于油菜根系腐解是个缓慢的过程,且需要消耗一定的N 素,导致拔节前土壤脲酶活性偏低;而到孕穗期,因为根系腐解,可能增加了土壤中的无机N、P含量,进一步促进了土壤脲酶活性的提高。
由图3-C可知,在0~100 cm 土层范围内,蔗糖酶活性随土层深度加深逐渐递减,在20~40 cm 至40~60 cm 土层下降最为显著,平均下降了45.32%。土壤蔗糖酶主要活跃在0~40 cm 土层,该土层土壤蔗糖酶活性占到了0~100 cm 土层蔗糖酶总活性的77.47%,这可能是由于在0~40 cm 土层中小麦根系分泌物质较多,土壤微生物较为活跃的原因;在3个处理之间,T在拔节期和孕穗期,其平均蔗糖酶活性均明显高于其他两组,这可能是由于油菜秸秆还田腐解后,为土壤带来了大量腐殖质,提高了土壤有机碳含量,进而促进了土壤蔗糖酶活性提高,保护了土壤蔗糖酶免遭分解,免遭变性;在不同生育期,孕穗期土壤蔗糖酶活性明显高于拔节期,说明孕穗期植株根际微生物更活跃,土壤中碳的固转速率更快。
由图3-D可知,孕穗期土壤含水量明显低于拔节期;0~60 cm 土层土壤含水量明显低于60 cm 以下土层;T含水量最高,CK0次之,CK1最低,但三者之间未达到显著差异(P>0.05,表略)。拔节期土壤平均含水量为13%,孕穗期明显降低,说明拔节期-孕穗期,作物生长量急速增加,为需水高峰期;0~60 cm 土层土壤含水量在9%~11%左右,60 cm 以下土层土壤含水量平均在13%以上,说明拔节期-孕穗期小麦根系的活跃吸收层主要在0~60 cm 土层,因此该层土壤含水量偏低,结合土壤脲酶活性发现,该层酶活性较高,也充分说明该层为活跃吸收层;而60 cm 以下土层土壤含水量较高,而脲酶活性较低,主要为生育后期深层根系吸收利用。此外,3个处理之间无显著差异(P>0.05,表略)。

图3 不同处理下小麦拔节期、孕穗期土壤有机质含量(A)、脲酶活性(B)、蔗糖酶活性(C)及土壤含水量(D)垂直分布比较Fig.3 Soil organic matter content(A),urease activity(B),surcease activity(C) and soil moisture (D) of vertical distribution comparison under the different treatments at jointing and booting stages of wheat
2.5 麦后复种饲料油菜对周年经济效益的影响
小麦价格以2.4元·kg-1计算,鲜油菜以160元·t-1计算;CK0农机费60元·666.7m-2,另外两组农机费用为90元·666.7m-2;种子费:油菜70元·kg-1(播种量1.5 kg·666.7m-2)、小麦2.4元·kg-1(播种量15 kg·666.7m-2);肥料投入:150元·666.7m-2。
由表6可知,CK0、CK1和T的年净收益分别为343.19、785.79和218.20元·666.7m-2,三者差异达到显著水平(P<0.05)。CK1的年净收益相较于CK0提高了128.97%,T 较CK0的年净收益降低了36.42%。可见,在夏休闲期复种饲料油菜且收获地上部分,可以明显提高年净收益。作为绿肥的饲料油菜还田处理,其年净收益虽不及CK0,但还田处理可以改善土壤质量,提高土壤可持续利用能力。

表6 不同处理对小麦周年经济效益比较Table6 Effect of different treatments on annual economic benefits
3 讨论
3.1 麦后复种饲料油菜对后作小麦植株生长的影响
水分是影响小麦生长发育的主要因素之一[15]。研究表明,夏休闲期种植油菜会消耗较多的土壤水分[16],导致后作小麦生长水分不足,土壤养分亏缺,从而影响小麦生长。本研究中,2个复种油菜处理(T和CK1)的小麦单株绿叶数在拔节初期后均显著高于不复种油菜处理(CK0),且T>CK1(P<0.05);复种油菜处理的小麦旗叶叶宽及叶面积也较高,表明在晋中地区,麦后复种饲料油菜有利于促进后作小麦分蘖发生,光合叶面积提高。气象资料表明(图4),2016年7-9月,试验地太谷县平均月降雨量为70.03 mm,水分充足,可以满足种植油菜需要消耗的土壤水分。此外,还田后的油菜,冬前的腐解过程,有利于后作小麦春季生长期植株N 素积累,进而提高干物质积累,提高产量。

图4 2015年7月-2016年7月太谷县的月平均降雨量Fig.4 The average rainfall per month in Taigu county from July,2015 to July,2016
3.2 麦后复种饲料油菜对后作小麦产量及品质的影响
单位面积总穗数、穗粒数、千粒重是评价小麦产量的重要指标[17]。本试验中,3个处理的总穗数和千粒重差异不显著(P>0.05),穗粒数则表现为T>CK0>CK1(P<0.05)。究其原因,单位面积总穗数取决于冬前茎蘖生长状况,试验3个处理的冬小麦植株在越冬期时株高、绿叶数、功能叶面积均无显著差异,可能是因为冬前植株苗小,其营养主要来源于土壤基础养分;另外,穗粒数主要取决于拔节期-孕穗期的小穗形成和小花分化,此生育期为小麦茎蘖春季生长高峰,需要吸收更多的土壤养分。本研究中,从起身期-孕穗期前,还田处理T的植株N 积累更高,说明T 处理土壤养分更充足,可能是由于此生育期气温升高,有利于还田油菜腐解,从而提高土壤养分,有利于植株吸收收用,进而促穗促花、保花增粒。此外,粒重形成期小麦植株所需的水分养分主要由深层土壤提供,而还田油菜主要在25~30 cm 耕层。植株N含量对小麦产量有着至关重要的影响[18],提高植株N含量是提高小麦产量的一种重要方式,绿肥可以转化土壤中的难溶性养分(如P)[19]。本试验中,小麦植株N 积累越冬期前表现为CK0明显高于CK1和T,返青期之后CK1尤其T的植株N 积累较高;花后20 d之前,T、CK1和CK0之间的单茎P含量无太大差异,但CK0略高;而花后20 d后,T的单茎P含量则不同程度地高于CK0和CK1。这可能是由于冬前油菜根茎叶腐解消耗了一部分土壤N、P 营养,而返青-灌浆期,腐解后的油菜提高并活化了土壤N、P 营养,表现为在N、P 吸收积累高峰期,植株N、P含量高。本试验中,T 明显提高了后作冬小麦理论产量,说明油菜秸秆还田对后作冬小麦产量的提升具有很大潜力。此外,T 也明显提高了后作小麦籽粒蛋白质总含量(P<0.05)。
3.3 麦后复种饲料油菜对后作小麦土壤肥力的影响
有机质含量高低能反映出土壤质量好坏[20]。本研究中,在拔节期和孕穗期,油菜全量还田均明显提高了土壤有机质含量,与前人研究结果相一致[21],这主要是由于油菜秸秆在腐解过程中,为土壤带来大量的氮源和碳源[22],增强了土壤中微生物及酶代谢[23]。从拔节期到孕穗期所有处理的土壤有机质含量均明显降低,这可能是由于此阶段为小麦生长高峰期,需要消耗大量土壤水分和养分。本研究结果还表明,油菜还田对0~40 cm 土层的土壤有机质含量的提高效果最好,且0~20 cm 土层的有机质含量提高最为显著,这是由于油菜秸秆主要翻压在土壤耕作层的原因,释放出的养分也大部分存在于土壤耕作层,因此对土壤耕作层的改良效果更好。
脲酶直接参与土壤含氮有机物的转化,其活性常用来反映土壤供氮水平[24]。本试验结果表明,土壤脲酶活性在10~20 cm 最高。根据吕宁等[25]的观点,脲酶主要来源于土壤微生物及植物活体分泌或植物残体释放。因此,其土壤脲酸活性高,一方面由于油菜秸秆的腐解提高了周围土层微生物的活性与数量,进而分泌更多的土壤脲酶;另一方面则是油菜秸秆残体释放了一部分脲酶。此外,CK1脲酶活性在拔节期低于CK0,而在孕穗期却高于CK0,这可能是由于油菜根系的腐解过程较为缓慢,需要一定的时间,其具体机理将在后续研究中进一步深入探讨。
土壤蔗糖酶可以通过酶解土壤中的大分子糖类参与有机碳的形态转化[26],同时也可以直接参与一些有机质的代谢,为土壤微生物提供能源[27],其常用来反映土壤熟化程度[28]。油菜秸秆中含有大量可溶性多糖、氨基酸以及大量的无机养分[29]。本研究中,油菜全量还田为蔗糖酶提供了更多的酶解底物,导致土壤蔗糖酶活性明显增加。同时,油菜全量还田增加了土壤有机质含量,也可保护蔗糖酶不受分解[30]。
综上所述,油菜全量还田对于后作小麦土壤肥力提升效果是最好的。
3.4 麦后复种饲料油菜对周年经济效益的影响
油菜-小麦周年经济效益取决于油菜种植带来的年净收益以及小麦生产带来的年净收益之和。本研究中,CK1年净收益较CK0提高了128.97%,而T年净收益最低,较CK0降低了57.28%。这可能是由于本研究是一年期试验,油菜改土效应促进后作小麦增产所带来的效益还不足以弥补或超过油菜栽培所消耗的资本投入。但油菜还田的改土效应已初见成效,后续将进行多年复种还田定位试验对改土效应与经济效益评价研究。
4 结论
在晋中地区,油菜还田可以促进后作小麦植株生长发育及干物质积累,从而提高小麦产量;还可以提高后作小麦籽粒蛋白质含量;同时促进土壤酶活性提高与有机质含量提升。此外,种植油菜且不还田可以提高周年经济效益。
