蛋氨酸二肽在动物生产中的应用研究进展
2020-03-13王卓铎孙广文张海涛
■刘 敏 王卓铎 孙广文 张海涛
(广东恒兴饲料实业股份有限公司 农业农村部华南水产与畜禽饲料重点实验室,广东湛江524000)
蛋氨酸(Methionine, Met),又名甲硫氨酸,具有旋光性,分为D和L两种构型。在机体内,只有L-Met能被直接利用,而D-Met 需先转化为L-Met,然后才能参与氨基酸代谢过程[1]。蛋氨酸是唯一的含硫必需氨基酸,参与胆碱、肾上腺素和肌酸的合成,磷的代谢和甲基的转移,以及为动物体内合成胱氨酸和蛋白质提供原料[2]。蛋氨酸是禽类玉米-豆粕型饲粮、奶牛泌乳的第一限制性氨基酸,是猪的第二或第三限制性氨基酸[3]。此外,蛋氨酸通常是水产动物饲料中的第一限制性氨基酸[4]。同时蛋氨酸是多种植物性蛋白原料的限制性氨基酸[5]。
饲料中使用的蛋氨酸产品主要有DL-蛋氨酸、晶体蛋氨酸、包膜蛋氨酸、蛋氨酸羟基类似物(Methio⁃nine hydroxy-analog,MHA)和蛋氨酸二肽(DL-methi⁃onyl-DL-methionine,Met-Met)等[6]。DL-蛋氨酸是一种D型和L型蛋氨酸各占50%的外消旋体氨基酸,主要通过化工合成[1]。晶体蛋氨酸是传统的化学合成蛋氨酸,其在水中的高溶失率导致水产饲料中外源补充晶体蛋氨酸效果不佳[7]。此外,晶体氨基酸能否为无胃鱼有效利用也存在争议,低投喂频率(1~2次/d)下可能存在不同步吸收的问题[8]。包膜氨基酸应用了包被及控释技术,可以有效缓解晶体蛋氨酸在水中的溶失并且解决了晶体氨基酸在肠道中与蛋白态氨基酸吸收不同步的问题。但包膜氨基酸的作用效果因水产动物种类而异,且包膜材料种类众多,价格不一,效果也不一致[7]。羟基蛋氨酸是以丙烯醛和甲硫醇为原料合成的,其生产工艺简单,原料用量少,副产物少,生产成本较低[9]。但目前生产中主要出现的缺点是有硫化氢气味。
Met-Met是一种使用化学合成方法得到的DL-蛋氨酸的二肽聚合物,室温下较稳定,不易溶于水,呈白色或棕色粉末状;组成Met-Met的混合物包括4种不同结构的二肽:DL-Met-Met、LD-Met-Met、DD-Met-Met和LL-Met-Met)[8,10]。Met-Met水溶性明显低于其他来源的蛋氨酸,因而其过胃效果明显,可避免在胃中被快速溶解。Met-Met过胃后经寡肽酶水解成2个单独的游离DL-Met,并缓慢释放,精确地用于蛋白质的合成,从而促进动物生长[11]。本文就Met-Met在畜禽动物和水产动物上的应用研究进展进行了综述,以期为蛋氨酸资源的节约利用,配方成本的降低,以及Met-Met在实际生产的应用提供理论依据,并为低鱼粉或无鱼粉饲料中蛋氨酸的添加与应用提供新的参考方向。
1 Met-Met在畜禽生产中的应用研究进展
1.1 Met-Met对禽类生长性能的影响
有关Met-Met对禽类生长性能影响的研究,目前仅见肉仔鸡中有报道。
鸡球虫病是由艾美耳科艾美耳属的七种球虫引起的鸡的寄生虫性疾病[12],可导致家禽肉和蛋生产出现严重损失。球虫在小肠内成倍增殖导致组织损伤,降低摄食率(FI)和饲料效率(FE)。随着自由散养家禽数量的增加,球虫病已成为极大的威胁。Souza等[13]探讨了艾美耳球虫感染和日粮中添加游离蛋氨酸或Met-Met对肉仔鸡生产性能的影响。14日龄的鸡经口腔分别接种堆型艾美耳球虫、早熟艾美耳球虫、巨型艾美耳球虫和缓艾美耳球虫的孢子囊。结果发现,与未经艾美耳球虫感染的肉仔鸡相比,感染了艾美耳球虫的肉仔鸡FI降低了13%,体增重(WG)降低了37%,饵料系数(FCR)升高了39%。且感染组的肉仔鸡法氏囊相对重量较低(51.8%),脾脏和肠相对重量较高(分别为53.6%和26.3%)。与无蛋氨酸添加的日粮饲喂的肉仔鸡相比,日粮中DL-Met和Met-Met的添加使得肉仔鸡WG升高了12%,FCR降低了12%。艾美耳球虫感染导致肉仔鸡空肠内发生炎症反应和氧化应激,进而可能影响肠上皮的完整性,最终导致肉仔鸡生长性能下降。而日粮中补充游离蛋氨酸和蛋氨酸二肽能缓解此炎症反应和氧化应激,从而提高肉仔鸡生长性能。
1.2 Met-Met对反刍动物生产性能的影响
目前有关Met-Met 对反刍动物生产性能影响的研究主要集中于其对奶牛泌乳的影响。
从营养角度讲,在满足奶牛乳腺总氮需要量的前提下,为奶牛乳腺提供乳蛋白合成所需的最佳氨基酸营养模式是提高奶牛乳蛋白水平的主要途径。为了保持氮和碳的摄取和排出平衡,乳腺组织必须摄取非游离态的氨基酸(小肽或蛋白质)来满足乳蛋白合成的需求[14]。奶牛乳腺上皮细胞对小肽的利用效率高于游离氨基酸,小肽转运体系的相对饱和性可能要比氨基酸转运体系高[15]。奶牛乳腺上皮细胞(BMECs)内乳蛋白主要包括:αs-酪蛋白(αs-CN)、β-酪蛋白(β-CN)、κ-酪蛋白、β-乳球蛋白、Ⅱ型小肽转运载体和氨肽酶氮[16]。
αs-CN是牛乳乳蛋白中含量最高的一种蛋白,占总蛋白的50%左右,因此用检测乳腺组织αs-CN 合成量来表示乳蛋白的合成有很强的代表性[17]。吴慧慧[17]研究了蛋氨酸及其二肽(Met-Met)不同组合对αs-CN合成的影响。试验共设5个组,各组的蛋氨酸浓度分别为50、67.5、85、102.5、120 μg/ml,Met-Met浓度分别为70、52.5、35、17.5、0 μg/ml。结果表明,Met 为85 μg/ml、Met-Met 为35 μg/ml 的组合中αs-CN 合成量最高,且显著高于其他各组。通过拟合曲线计算Met 浓度为81 μg/ml、Met-Met浓度为38 μg/ml时αs-CN合成量最高。该研究同时得出,在以二肽部分替代游离氨基酸的条件下,Lys:Met:Met-Met:Thr:Phe 为100:22.5:1.5:56:63 时,αs-CN 合成量最高,且高于完全以游离氨基酸配比时的αs-CN的合成量。
Wang等[18]探讨了Met-Met在β-CN合成中的转运特性及利用。试验研究了时间、pH 值、浓度、温度和抑制剂对BMECs 中的异硫氰酸荧光素蛋氨酸二肽(Fluorescein isothiocyanate Met-Met,Met-Met-FITC)传输特性的影响;Met-Met 的浓度分别为0、20、40、80、120 μg/ml 和160 μg/ml;抑制剂为Janus 激酶2(JAK2,50 μmol/l)和雷帕霉素(mTOR,100 ng/ml)。结果表明,BMECs 对Met-Met-FITC 的吸收在前15 min很快,15 min 后饱和;Met-Met-FITC 的摄取在pH 值6.5时达到峰值;Met-Met浓度依赖性地增加了BMECs中肽转运蛋白2(PepT2)的表达和β-CN 的合成,且最优浓度为80 μg/ml;JAK2和mTOR的抑制显著降低了经Met-Met 诱导的BMECs 的细胞活力和β-CN 的合成。总之,Met-Met经PepT2吸收后,通过激活BMECs中的JAK2-STAT5和mTOR信号通路来增强细胞增殖并促进β-CN合成。
1.3 Met-Met对畜禽抗氧化性能的影响
Souza 等[13]的研究发现,肉仔鸡接种艾美耳球虫孢子囊144 h后,肉鸡空肠中氧化物水平升高,如亚硝酸盐和硫代巴比妥酸反应物(TBARs)升高,羰基化蛋白含量(CP)较高,抗氧化性能(TAC)和过氧化氢酶(CAT)活性较低,艾美耳球虫感染导致肉仔鸡空肠内发生炎症反应和氧化应激。而日粮中添加游离蛋氨酸和Met-Met,使得受感染的肉鸡TAC 升高,保护了肉仔鸡肠道细胞免受球虫感染引起的氧化损伤。
1.4 Met-Met对畜禽相关基因表达的影响
Souza等[13]的研究同时发现,Met-Met可影响肉仔鸡免疫应答基因的表达。该研究推测,艾美耳球虫感染抑制了过氧化氢酶(CAT)基因和超氧化物歧化酶1(SOD1)基因的表达;并同时抑制了中性氨基酸转运体1(B0AT1)、肽转运体1(PEPT1)、Toll样受体5(TLR5)、白细胞介素2(IL2)和闭合蛋白(OCLN)的基因表达,增强了阳离子氨基酸转运体1(CAT-1)和干扰素γ(IFNG)的基因表达。而DL-Met和Met-Met的添加改变了这一现象,且日粮中添加Met-Met的肉鸡PEPT1表达水平最高,无添加组的肉鸡TLR5表达水平最低。
相关研究表明,Met-Met 亦可对奶牛泌乳进行分子水平的调控。在体外培养体系中添加Met-Met 和Lys-Met二肽,比添加相同含量的游离氨基酸更能促进αs1-CN基因的表达。改变培养液中赖氨酸和蛋氨酸的比例能影响αs1-CN基因的表达,当两者比例在3.5:1时,αs1-CN基因表达最强[15]。此外,杨金勇等[19]的研究表明,添加蛋氨酸浓度为57 μg/ml时αs1-CN基因的表达最强。用60 μg/ml Met-Met 替代等量的蛋氨酸时,Met-Met 组αs1-CN基因表达极显著高于蛋氨酸组(P<0.01)。陈璐等[16]以不同含蛋氨酸二肽(P-Met-Met、PMet-Lys、P-Met-Trp、P-Met-Phe、P-Met-Thr、P-Met-Ile、P-Met-Leu、P-Met-Val)等量替代对应的游离氨基酸,结果发现含蛋氨酸二肽均能促进BMECs内乳蛋白合成相关基因的表达,其中尤以P-Met-Met的效果最好。高学军等[20]与毕微微等[21]的研究也都表明,Met-Met 可以促进奶牛乳腺上皮细胞增殖并促进β-CN 基因的表达,进而影响乳腺发育及泌乳等过程。
2 Met-Met在水产养殖动物中的应用研究进展
2.1 Met-Met对水产养殖动物生长性能的影响
相较于其他蛋氨酸源,Met-Met 具有的水溶性低、不占用单体氨基酸吸收通道、缓释等优良特性使其在水产动物上的研究逐渐增多。有关Met-Met 在草鱼幼鱼[22]、幼鲤[8]、凡纳滨对虾[23-24]、真鲷[25]上的研究均表明,日粮中添Met-Met可显著改善这些养殖动物的生长性能。然而,日粮中添加Met-Met对牛蛙的生长性能无显著影响[26]。
苏玥宁[22]分别以含2.50 g/kg DL-Met的日粮和不同水平Met-Met(0、0.79、1.44、1.84、2.22 g/kg和2.85 g/kg)的日粮饲喂初重(10.10±0.03) g的健康草鱼10周,考察Met-Met对其生产性能的影响。结果发现,在蛋氨酸缺乏的日粮中添加适量的Met-Met提高了幼草鱼增重百分比(PWG)、特定生长率(SGR)、FI和FE,升高了肌肉和肝胰脏中谷草转氨酶(GOT)和谷丙转氨酶(GPT)的活力,并降低了血浆氨含量(PAC)(P<0.05)[22]。尹子煜等[8]通过在低蛋氨酸饲料中添加不同水平的Met-Met,饲喂初重(16.7±0.08) g的幼鲤,以评价Met-Met在幼鲤饲料中的有效性。结果表明,与未添加组相比,各Met-Met添加组幼鲤的末均重、增重率、SGR均显著增加(P<0.05),而FCR均显著降低(P<0.05)。Mamauag等[25]比较了两种分子形式的蛋氨酸(DL-Met和Met-Met)对真鲷仔稚鱼(体重42 mg)和幼鱼(体重0.75 g)生长效果的影响。结果表明,DL-Met和Met-Met可以被真鲷仔稚鱼和幼鱼同等利用,且对氨基酸进行包被显著改善了仔稚鱼和幼鱼的生长性能。
Wang等[23]探讨了植物性蛋白饲料中添加不同水平的Met-Met对凡纳滨对虾生长性能的影响,试验为期10周。阳性对照组(PC)含20%的鱼粉,阴性对照组(NC)含8%的鱼粉,其他六个试验组在NC的基础上分别添加不同水平(0.05%、0.1%、0.15%、0.20%、0.25%、0.30%)的Met-Met,虾初重(0.98±0.02) g。结果发现,鱼粉水平从20%到8%,并添加不同水平的Met-Met,对凡纳滨对虾存活率(SR)、FE、蛋白质效率比(PER)、全虾和肌肉组成没有显著影响(P>0.05)。然而,饲料Met-Met 添加量≤0.20%时,WG和SGR随Met-Met添加水平的升高而显著增加;Met-Met添加量继续升高,WG和SGR趋于平稳状态。Xie等[24]也研究了降低日粮鱼粉水平同时添加蛋氨酸对凡纳滨对虾的影响。结果表明,鱼粉水平从260 g/kg 降至100 g/kg 对凡纳滨对虾的SR、FCR、PER或蛋白质保留率(PR)无显著影响。通过在鱼粉含量为100 g/kg的饲料中增加DL-Met的水平,对虾的生长性能(FBW、WG和SGR)显著提高(P<0.05)。鱼粉水平为100 g/kg的饲料中额外添加3.0 g/kg的DL-Met或1.0 g/kg的Met-Met也能与阳性对照组达到同样效果。DL-Met(3.0 g/kg)、Met-Met(1.0 g/kg)组获得了最高的生长性能(FBW、WG和SGR),且显著高于无蛋氨酸添加组(P<0.05),但与全鱼粉组(260 g/kg)、DL-Met(1.0 g/kg)、DL-Met(2.0 g/kg)组间无显著性差异(P>0.05)[24]。
冯伟[26]以鱼粉、豆粕和大豆分离蛋白为蛋白源,磷脂油、鱼油和豆油为主要脂肪源,面粉为主要糖源,配制粗蛋白水平为40%、脂肪水平为6%的基础饲料,在基础饲料中分别添加0、0.4%、1.2%和2.0%的Met-Met,设计9种等氮等能的实验饲料,饲喂初重(40.01±0.24) g 的牛蛙8 周。结果表明,添加Met-Met 组与其他各试验组的牛蛙WGR、PER、PD(蛋白质沉积率)、FCR、FI和SR之间均无显著差异(P>0.05)。
2.2 Met-Met对水产养殖动物免疫性能的影响
头肾和脾脏是鱼类主要的免疫器官[27],肠道、皮肤和鳃是鱼类重要的黏膜免疫组织[28]。鱼类的免疫功能与其抗菌能力和调节炎症反应的能力密切相关[29]。
Su 等[30]探讨了日粮中添加Met-Met 对草鱼幼鱼肠道、皮肤、鳃和免疫器官的免疫功能和潜在信号分子的影响。其研究发现,Met-Met 增强了草鱼幼鱼肠道溶菌酶(LZM)和酸性磷酸酶(ACP)的活性,提高了补体(C3 和C4)和免疫球蛋白M(IgM)的含量。Met-Met 增强草鱼肠道、鳃、头肾、脾脏和皮肤免疫功能可能与其提高抗菌物质活力和含量,以及通过NF-κB信号途径下调促炎细胞因子和TOR 信号途径上调抗炎细胞因子基因表达,降低炎症反应有关。
2.3 Met-Met 对水产养殖动物形体指标和体成分的影响
相关研究表明,饲料中添加Met-Met可显著影响幼鲤[8]、凡纳滨对虾[23-24]部分形体指标和体组成,对牛蛙的形体指标和体组成则无显著影响[26]。尹子煜等[8]的研究发现,与未添加Met-Met 的试验组相比,低蛋氨酸饲料中添加不同水平Met-Met 后幼鲤的肝胰指数和脏体指数均有所降低;全鱼水分含量增加;粗灰分含量显著降低(P<0.05);全鱼中Met、Met+Cys、Lys、Trp、Ile、Leu、Val、His 的含量显著增加(P<0.05)。鱼体PD随饲料Met-Met添加水平的增加先增加后降低,饲料Met含量为7.6 g/kg时,PD达到最大;必需氨基酸沉积率显著升高(P<0.05)。Wang 等[23]报道,在植物性蛋白饲料(阴性对照组含8%鱼粉)中添加不同水平的Met-Met,各添加组的虾肌肉中总必需氨基酸(EAA)含量显著高于阴性对照组(P<0.05)。Xie 等[24]的研究发现,鱼粉含量为100 g/kg的饲料中添加Met-Met,虾的EAA 含量与阳性对照组(鱼粉含量为260 g/kg)的虾的EAA无显著性差异(P>0.05),但虾的全身和肌肉蛋白含量显著低于阳性对照组(P<0.05)。然而,冯伟[26]的研究发现,Met-Met添加组与未添加组相比,牛蛙全体水分、粗蛋白和粗灰分、牛蛙脊椎骨和腿骨粗灰分之间无显著差异(P>0.05),同时实验各组牛蛙肌肉水分、粗蛋白、粗灰分和氨基酸组成亦无显著差异(P>0.05)。
2.4 Met-Met 对水产养殖动物血清和肝脏生化指标的影响
血液是动物体内循环系统的重要组成部分,起着物质运输、生理调节及生理防御等重要作用[31]。血清能够为机体提供营养物质、激素和生长因子等,血清生化指标可反映机体的健康状况[32]。冯伟[26]关于牛蛙饲料中添加Met-Met的研究发现,随着饲料添加Met-Met 水平的升高,实验各组血清甘油三酯(TG)、总胆固醇(T-CHO)、葡萄糖(GLU)和GPT之间无显著差异(P>0.05),而Met-Met 添加量为2.0%的试验组牛蛙血清GOT 显著低于Met-Met 添加量为0.4%的试验组和Met-Met 添加量为1.2%的试验组(P<0.05)。饲料中添加蛋氨酸或Met-Met 对实验各组牛蛙肝脏超氧化物歧化酶(SOD)、丙二醛(MDA)、CAT、GOT和GPT均无显著影响(P>0.05)。总之,饲料不同蛋氨酸或Met-Met水平可显著影响牛蛙部分血清生化指标,适宜水平的蛋氨酸可降低牛蛙血清T-CHO的含量。尹子煜等[8]报道,低蛋氨酸饲料中添加4.0 g/kg 的Met-Met,幼鲤血浆高密度脂蛋白胆固醇(HDL-C)/T-CHO显著高于FM组(10%鱼粉和3.0 g/kg晶体DL-Met)和MO组(0 g/kg的Met-Met)(P<0.05),适宜添加量的Met-Met 可有效促进幼鲤脂肪的转运与代谢,提高其抗氧化能力。
2.5 Met-Met对水产养殖动物相关基因表达的影响
研究表明,Met-Met 可对水产养殖动物的生长和免疫性能进行分子水平的调控。胰岛素样生长因子(Insulin like growth factor,IGF)是一类多功能细胞增殖调控因子,在细胞的分化、增殖、个体的生长发育中具有重要的促进作用,同时也是生长激素启动生长活性的主要介导者[33]。IGF-I不仅可以作为鱼类生长的重要标志,也同样可以作为评价水产动物营养状况的指标之一[34]。冯伟[26]的研究表明,随着牛蛙日粮中Met-Met 添加水平的升高,各组之间牛蛙肝脏生长发育相关基因IGF-I 和IGF-IR 基因的mRNA 呈上升趋势,但各添加浓度间无显著性差异(P>0.05)。Met-Met(2.0 g/kg)组IGF-I 基因的mRNA 表达量显著高于Met(2.0 g/kg)组(P<0.05);Met(0.4 g/kg)组和Met(1.2 g/kg)组IGF-IR 基因的mRNA 分别显著高于Met-Met(0.4 g/kg)组和Met-Met(1.2 g/kg)组(P<0.05)。饲料不同Met或Met-Met水平可显著影响肝脏生长相关基因的表达,蛋氨酸水平过低或过高均会下调IGF-I 和IGF-IR mRNA 的表达。此外,Met-Met 提高草鱼肠道、鳃、头肾、脾脏和皮肤结构完整性可能与其通过Nrf2信号上调抗氧化酶基因表达并提高酶活力,增强抗氧化损伤能力;通过下调促凋亡蛋白,上调抗凋亡蛋白基因表达,抑制细胞凋亡;以及上调细胞间紧密连接蛋白基因表达有关[22,30]。
2.6 Met-Met在水产料中的适宜添加量
相关研究基于不同的试验条件下Met-Met 对不同的水产养殖动物养殖效果的分析,得出了饲料中对应的Met-Met适宜添加量,如表1。
研究表明,基于PWG、肠炎发病的预防和免疫指数(LZM活性)考虑,得出草鱼幼鱼(10.10~126.39 g)饲料中Met-Met的最适添加量为1.61、1.64 g/kg和1.68 g/kg,对应的基础日粮总含硫氨基酸(TSAA)含量为8.03 g/kg,其中蛋氨酸4.26 g/kg,半胱氨酸3.77 g/kg[22,30]。在Met缺乏的基础饲料(含4.8 g/kg 的Met)中添加适量Met-Met可有效促进幼鲤摄食和生长,促进脂肪的转运与代谢,提高抗氧化能力。基于WG、FCR和PD的回归分析表明,幼鲤Met需求量为7.6~8.1 g/kg(饲料中Met-Met添加量为2.95~3.47 g/kg),占饲料蛋白质的2.0%~2.1%[8]。Xie等[24]的研究表明,鱼粉水平为100 g/kg的凡纳滨对虾饲料中额外添加3.0 g/kg 的DL-Met 或1.0 g/kg 的Met-Met能与鱼粉水平为260 g/kg的饲料(不额外补充氨基酸)达到同样的效果。此外,在低鱼粉日粮中添加限制性氨基酸蛋氨酸,似乎可以提高消化蛋白水解酶活性,提高干物质和蛋白质的消化率,最终在日粮鱼粉水平降低的条件下促进凡纳滨对虾的生长。Wang等[23]的研究基于折线模型分析显示,以蛋氨酸二肽为蛋氨酸源时,凡纳滨对虾对蛋氨酸的需求量为0.87%。
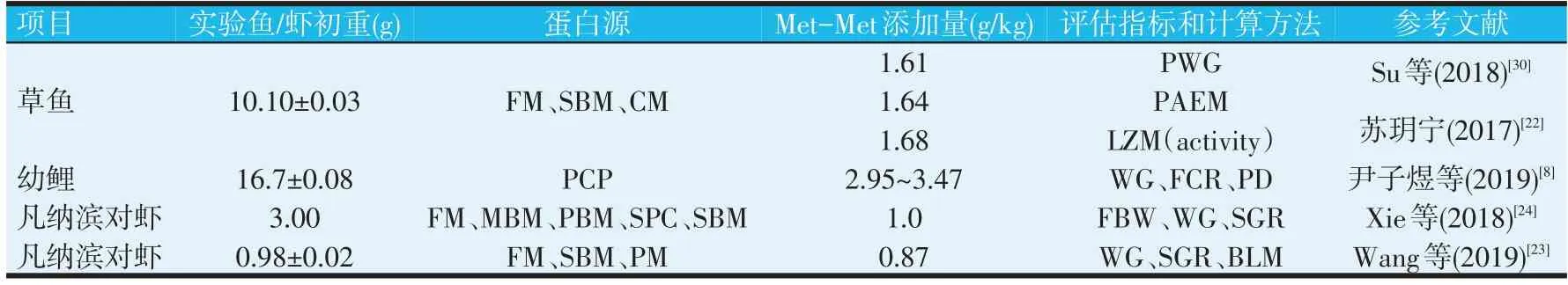
表1 水产料中的蛋氨酸二肽需求
3 Met-Met与DL-Met的生物利用率的比较
Santos等[35-36]进行了Met-Met与Met在断奶仔猪和生长猪体内的生物利用度(RBV)的比较。结果发现,添加DL-Met或DL-Met-Met后,尿氮排泄量呈直线下降(P≤0.01;线性),氮保留量、氮滞留量呈直线上升(P≤0.02;线性)。基于线性回归模型分析发现,等摩尔的DL-Met-Met 相 对DL-Met 的RBV 为:氮 保 留 量111%(95% CI:63%~158%),氮(%摄入)滞留量109%(95% CI:57%~160%),氮(%吸收)滞留量98%(95%CI:43%~154%)。且添加DL-Met 或DL-Met-Met 后,总平均日增重(ADG)和增重:饲料(G:F)呈线性增加(P<0.01)。对于ADG,等摩尔基础上以斜率比估算,Met-Met 的RBV 为104%(95% CI:66%~141%);对于G:F,等摩尔基础上,Met-Met 的RBV 为117%(95%CI:61%~174%)。因此,Met-Met可作为断奶仔猪和生长猪的蛋氨酸来源,且对猪来说Met-Met与DL-Met的生物利用度没有差异。杨金勇等[19]的研究也表明,奶牛乳腺可以利用Met-Met,而且其利用效率高于蛋氨酸。
Niu等[37]比较了Met-Met(AQUAVI®) 与DL-Met在凡纳滨对虾幼虾上的利用效果。结果基于WG、SGR、FCR的非线性回归模型分析表明,AQUAVI®Met-Met比DL-Met具有更高的RBV(P<0.05),且相对值分别为285.8%、276.4%和300.4%。此外,不同蛋氨酸源的RBV也存在差异,且AQUAVI®Met-Met比DL-Met更适用于凡纳滨对虾。此外,Wang 等[23]的研究发现,凡纳滨对虾饲料中添加Met-Met显著提高了干物质、粗蛋白、脂类、磷和EAA的表观消化率(P<0.05)。Xie等[24]在凡纳滨对虾上的研究得到了类似的结果,相比于阳性对照组与DL-Met添加组,Met-Met组的干物质和粗蛋白的表观消化率最高(P<0.05);且Met-Met组的虾肠道蛋白水解酶活性最高(P<0.05)。Mamauag等[25]的研究也表明,DL-Met和Met-Met可以被真鲷仔稚鱼和幼鱼同等利用。Su等[30]报道,添加Met-Met可改善鱼类的生长和肠道免疫功能,且在生长性能的提高和肠道免疫功能的增强方面,最佳的Met-Met添加量效果优于最佳的DL-Met添加量。饲料中Met-Met所提供的Met与天然蛋白质源中Met在消化吸收与利用上特性相似,以Met-Met为来源评估幼鲤Met需求量会更加准确。
4 小结与展望
综上,饲料中添加Met-Met 可促进奶牛泌乳;可提高肉仔鸡、草鱼幼鱼、凡纳滨对虾、真鲷仔稚鱼的生长性能、抗氧化性能和免疫性能,但对牛蛙的相关性能没有显著性影响;Met-Met 可对肉仔鸡和草鱼的免疫反应、牛蛙肝脏的生长发育以及奶牛泌乳进行分子水平的调控;且饲料中添加Met-Met 对草鱼幼鱼、凡纳滨对虾、真鲷仔稚鱼的体组成有显著性影响。在断奶仔猪和生长猪上的研究表明,Met-Met 与DL-Met的生物利用度没有差异;而在凡纳滨对虾、草鱼幼鱼、幼鲤上的研究表明,Met-Met 的生物利用度优于DLMet 的生物利用度,且饲料中添加Met-Met 提高了干物质和粗蛋白的表观消化率。
由此可见,Met-Met 在畜禽和水产动物的生长效果上均有优良表现,且Met-Met相较于其他蛋氨酸源更稳定,水溶性低,缓释,减少了对单体蛋氨酸的消耗,降低了饲料成本,且因过胃效果明显减少了被代谢造成的浪费,从而降低了对环境的污染和含氮废物的排放,有助于水质改良和环境保护。Met-Met 的开发和利用研究,将为饲料工业中氨基酸的应用提供新思路,对饲用氨基酸产业意义重大。
