东海北部小型底栖动物群落对径流及黑潮暖流入侵的响应
2020-03-12刘清河马林李新正
刘清河,马林,李新正,3,4*
( 1. 中国科学院海洋研究所,山东 青岛 266071;2. 中国科学院大学,北京 100049;3. 中国科学院海洋大科学中心,山东 青岛 266071;4. 青岛海洋科学与技术国家试点实验室 海洋生物学与生物技术功能实验室,山东 青岛 266237)
1 引言
海洋底栖生物是海洋生态系统中种类最多、生态关系最为复杂的类群,其在海洋生态系统的物质循环和能量流动中具有举足轻重的作用[1]。小型底栖动物作为底栖动物中的重要组成类群,是底栖食物链/网的重要组成环节,也是仔、稚幼鱼的优质开口饵料以及良好的环境指示生物[2]。虽然小型底栖动物的个体较小,但其丰度值较高,在特定环境中,如河口软泥地区,其生物量甚至可以和大型底栖动物持平[3]。相关研究还表明,小型底栖动物在刺激微生物代谢、加速生源要素矿化等方面具有重要的作用[4–5],因此小型底栖动物在底栖生态系统的生物地化循环和能量代谢中具有十分重要的作用。
东海是西太平洋重要的边缘海以其广阔的大陆架和复杂的环流系统著称,包括闽浙沿岸流、台湾暖流、黑潮以及长江冲淡水等水系[6–8]。受各种水系综合作用的影响,东海北部特别是长江口及其外海区域生态系统的物质交换和能量流动频繁,对人类活动影响和环境变化更加敏感。近年来,人类活动对长江口及邻近海域的影响极为显著,水体富营养化、重金属污染、过度捕捞等问题严重,以及海洋酸化和气候变暖等环境威胁,导致该海区赤潮灾害频发[9],低氧海区持续扩大[10],渔业资源减少,鱼类多样性下降[11–12],以及大规模水母爆发等[13–14]。因此,探明该海域生态灾害发生的原因和机理、揭示不同生态类群对环境变化的响应,对保护生态系统健康稳定发展和预防未来潜在生态灾害等问题具有十分重要的意义。
目前有关东海及长江口海域底栖生物调查主要以大型底栖动物为主,大型底栖动物群落分布规律以及其和环境要素的相关关系也被逐步揭示,例如大型底栖动物群落结构和多样性[15],大型底栖动物的长期变化和季节变化[16–17],黑潮对大型底栖动物分布的影响[18],底栖动物群落变化与全球气候变化的联系等[19]。有关小型底栖动物在东海北部及长江口海域的研究内容主要集中于小型底栖动物丰度和生物量以及海洋线虫的群落结构上[20–24]。有关小型底栖动物和环境因子的相关性研究不够透彻,例如对低氧的响应,黑潮对其分布的影响等。因此,针对以上问题,本文对东海北部及近长江口海域小型底栖动物和环境因子进行了取样调查,以揭示小型底栖动物在该海域的分布规律以及环境因子对其影响,探究小型底栖动物对特殊环境的响应等,以期为研究该海域底栖生态系统中小型底栖动物提供数据参考,进一步提高人们对黑潮对底栖动物群落影响的认识,探究其对环境变化的响应机制和保护海洋生态系统提供科学依据。
2 材料与方法
2.1 研究海域和站位
分别于2016 年9 月9−12 日和12 月12−18 日,搭载“科学三号”科学考察船,对位于28°~33°N 和122°~125°E 海域间的20 个采样站进行了采样调查,采样站位见图1。
2.2 样品采集和处理
采用0.1 m2箱式采泥器获取底层沉积物样品。为保证表层沉积物的完整性,采用塑料软管将上覆水抽离并过筛。然后迅速使用内径为2.9 cm 的有机塑料取样管,从表面未受扰动的沉积物中取3 个重复样。每个芯样只取8 cm,取出后立即按0~2 cm、2~5 cm和5~8 cm 分层装瓶,加入10%甲醛溶液固定,轻轻震荡,使沉积物充分分散开。此外,另采集部分沉积物样品,冷冻保存(−20℃),用于实验分析用,测定沉积物各参数指标。水层环境因子的测定使用随船温盐深测定仪(CTD)进行现场测定。
沉积物有机质含量、重金属含量以及粒度组成均由中国科学院海洋研究所分析测试中测定。
2.3 数据处理和分析
生物量换算:对于小型底栖动物生物量的计算,目前普遍采用小型底栖动物各类群干重系数乘以相对应各类群的平均丰度值的方法计算获得,其单位为μg/(10cm2)。本研究中小型底栖动物各类群干重参照Widbom[25]和刘清河等[26],桡足类的平均个体干重参照McIntyre[27]的研究结果。
数据分析:利用PRIMER 6.0 & PERMANOVA+软件包[28]对小型底栖动物群类结构进行等级聚类(CLUSTER)分析,MDS 排序和相似性分析(ANOSIM)。所测环境因子和小型底栖动物之间的相关性分析使用SPSS 19.0 进行分析。利用PRIMER 软件包中的BIOENV 分析确定影响小型底栖动物群落组成的重要环境要素。
3 结果
3.1 研究海域环境特征
3.1.1 水深、底层水温和盐度
水深:根据随船CTD 现场实测结果,9 月采样站位平均水深为46.5 m,D1 站位水深最浅(21 m),D6站位最深(66 m);12 月采样站位平均水深58.3 m,A1 站位水深最浅(25 m),BS 站位最深(79 m)。两航次水深均表现出由近岸向外海逐步增加的趋势,相较于长江口附近,浙江外海水深显著增加。

图 1 采样站位Fig. 1 Sampling sites
水温:9 月航次各站位平均底层水温为21.4℃,C6 站位底层水温最低(14.2℃),C1 站位底层水温最高(24.3℃);平均表层水温为26.1℃,D8 站位表层水温最高(27.6℃),C1 站位表层水温最低(25.3℃);整体上,表层水温高于底层水温,除C6 站位外,底层水温随深度的增加并无显著变化。12 月航次各站位平均底层水温19.4℃,A1 站位底层水温最低(16.2℃),A6−AS 站位均表现出较高的底层水温(20.3℃);各站位平均表层水温为19.3℃,和底层平均水温持平,A1 站位水温最低(15.7℃),A6−AS 站位表层水温最高(20.3℃);各站位表层水温和底层水温基本保持一致,由近岸到外海水温略有增加,近岸底层略高于表层水温。
盐度:9 月航次平均底层盐度为32.9,C 断面底层盐度变化不大,D 断面长江口附近盐度明显低于外海盐度;平均表层盐度为29.4,显著低于底层盐度,在D 断面由长江口向外海盐度逐渐增加。12 月航次,平均底层盐度为33.5,除A1、B1 两个站位盐度略低以外,其他站位底层盐度基本一致;平均表层盐度为32.8,低于平均底层盐度,A1、B1 两站位表层盐度显著低于其他站位,各站位表层盐度从近岸向外海略微增加(各站位水深和温盐数值见表1)。
3.1.2 沉积物粒度组成、有机质含量、重金属含量
粒度组成:在粒度组成上,研究海域沉积物主要以粉砂(4~63 μm)和细砂(63~250 μm)为主,其次是黏土(<4 μm),中砂及以上颗粒(>250 μm)含量极少;其中C1、C5、D1、A1、A5、B1、B2 和B3 站位粉砂含量较高,均超过70%(图2),并且均靠近近岸区,除C5 站位以外。各站位平均粒径差异较大(图3),最小平均粒径为13.38 μm(B2 站位),最大值为153.25 μm(B5 站位),沉积物平均粒径大小和粉砂含量呈显著负相关(p<0.01)。研究海域各站位沉积物分选程度较差(1<σi<2),超过50%的站位分选程度差(分选系数σi>2),沉积物偏态(SK)很正偏(SK>0.3),频率正态分布曲线峰值尖锐(KG>1.56)。整体上,研究海域从近岸向外海,沉积物组成主要有粉砂质向砂质转变,粒径逐渐变粗,分选程度变差。
有机质含量:9 月航次各站位平均有机质含量为0.39%,其中D1 站位含量最高为0.56%;12 月航次各站位平均有机质含量为0.53%,其中A5 站位含量最高为0.79%。整体上,近岸有机质含量略高于外海,表现出陆源输入的影响,这一特征在B 断面更为显著。
重金属含量:对研究海域沉积物中6 种常见重金属进行了测定分析。依照GB 18668–2002《海洋沉积物质量》标准,沉积物中Pb、Zn 和As 含量均符合一类标准要求,Cr 和Cu 含量在近岸站位含量超过一类标准,Cd 含量超标严重且分布不均匀,在A1 站位高达3.9 mg/kg,而在部分站位Cd 含量为0。和沉积物有机质含量分布规律近似,研究海域重金属含量表现出陆源输入的影响,近岸站位重金属含量略高于外海站位(表2)。

表 1 各站位水深、底层和表层水温和盐度Table 1 Depth, temperature and salinity of the surface and bottom water of each site
3.1.3 环境因子分析
利用SPSS19.0 对研究海域各种环境因子进行相关性分析(表3),结果表明:底层水盐度、沉积物砂的含量、沉积物平均粒径与水深呈显著正相关(p<0.01);沉积物粉砂含量、重金属含量(Cd 除外)与水深呈显著负相关;沉积物有机质含量、重金属含量(Cd 除外)与粉砂和黏土含量呈显著正相关(p<0.01)与砂含量和平均粒径呈显著负相关(p<0.01);各重金属含量之间存在显著的正相关性(Cd 除外)。

图 2 各站位沉积物不同粒径颗粒所占百分比Fig. 2 Percentages of different grain sizes of the sediment in each site

图 3 各站位沉积物颗粒平均粒径Fig. 3 Average grain size of the sediment of each site

表 2 沉积物有机质和部分重金属含量Table 2 Organic matter and heavy metal contents of the sediment
利用PRIMER6.0 对水深(D)、盐度(S)、温度(T)、有机质含量(TOC)、重金属含量、黏土含量(Clay)、粉砂含量(Silt)、砂含量(Sand)等13 种环境因子进行PCA 分析(图4)结果表明:主成分轴1(PC1)可解释环境变异度的66.9%,主成分轴2(PC2)和主成分轴1(PC1)累积可解释环境变异度的80.9%;对PC1 贡献较大的主要是沉积物重金属(Cd 除外)、粉砂、砂以及有机质含量,从左向右表示沉积物中重金属和粉砂含量减少,砂含量增多;对第PC2 影响较大的因素依主要是沉积物Cd 含量、盐度、温度和水深,从下向上表示水深、盐度和温度逐渐降低,沉积物Cd 含量增加。
3.2 小型底栖动物
3.2.1 小型底栖动物类群组成、丰度和生物量
研究海域共鉴定出16 个小型底栖动物类群(C3 站位未做定量研究),9 月航次16 个类群,12 月航次13 个类群,包括自由生活海洋线虫、底栖桡足类、动吻类、多毛类、双壳幼体、介形类、涡虫、端足类、海螨类、异足类、蛇尾幼体、星虫类、枝角类、等足类以及涟虫类,其中海洋线虫是绝对优势类群,其次为底栖桡足类、动吻类、多毛类以及其他类群。9 月航次小型底栖动物平均丰度为(1 758±759)个/(10cm2),其中海洋线虫占总丰度的95.6%,桡足类1.57%,动吻类1.06%;平均生物量为(1 216.4±464.7)μg/(10cm2)(干重),其中线虫占55.26%,介形类18.19%,多毛类15.11%,桡足类4.21%,动吻类3.07%;12 月航次平均丰度为(2 011±1 471)个/(10 cm2),线虫占95.6%,桡足类2.03%,动吻类1.51%;平均生物量为(1 143.0±755.0)μg/(10 cm2)(干重),线虫占67.28%,多毛类12.21%,桡足类6.64%,介形类6.41%,动吻类5.30%。
两航次相比较,9 月航次小型底栖动物类群多于12 月航次,但在丰度和生物量上均小于12 月航次。其中9 月航次D1 站位小型底栖动物丰度最高,为(2 970±1 272)个/(10 cm2),C1 站位丰度最低为(929±245)个/(10 cm2),其生物量也最低(473.0±193.3)μg/(10 cm2)(干重),但生物量最高值出现在D6 站位(1 870.4±369.0)μg/(10 cm2)(干重),高于D1 站位,各站位间小型底栖动物丰度差异不显著;12 月航次中,B3 站位丰度和生物量最高分别为(4 837±1 415)个/(10 cm2)和(2 648.1±713.1)μg/(10 cm2)(干重),BS 站位丰度和生物量均最低分别为(269±128)个/(10c m2)和(217.9±80.9)μg/(10 cm2)(干重)。在水平分布上,不同海域间小型底栖动物丰度差异显著(p<0.01),近岸站位小型底栖丰度显著高于外海站位,在整体上,两航次间小型底栖动物总丰度无显著差异,除涡虫外,各个类群在丰度组成上无显著差异(图5)。

表 3 研究海域环境因子间相关性Table 3 Correlation between environmental variables of the study area
3.2.2 小型底栖动物群落结构

图 4 研究海域环境因子主成分分析Fig. 4 Results of PCA of environmental variables of the study area

图 5 小型底栖动物丰度和生物量分布Fig. 5 Distribution patterns of meiofaunal abundance and biomass
对研究海域小型底栖动物丰度进行CLUSTER 聚类分析,结果表明:在9 月航次,各站位小型底栖动物的类群组成在82%相似水平上可以划分为两组,C1、C5、D4 站位划分为一组,剩余站位划分为另外一组,SIMPER 分析结果显示,线虫、桡足和动吻类是影响两组差异的主要类群,累计解释总变异度的68.5%;12 月航次,各站位小型底栖动物的类群组成在78%相似水平上可以划分为3 组,组Ⅰ只包含BS 一个站位,组II 包括A1、A5、B1、B2、B3 站位,组Ⅲ包括A6、A8、A9、AS、B4、B5 站位,其中组Ⅱ主要为近岸站位,组III 主要为外海站位,ANOSIM 检验结果表明组Ⅱ和组Ⅲ之间差异显著(R=0.672,p<0.01),SIMPER 分析结果表明线虫、动吻类和桡足类是影响两组差异的主要类群,累计解释总变异度的83.21%;两航次所有站位小型底栖动物的类群组成在78%相似水平上主要可以划分为两组(BS 站位除外,图6),SIMPER 分析结果显示影响组间差异的主要类群为线虫、动吻、桡足、介形和多毛类,累计解释总变异度的84.33%;对两航次间小型底栖动物群落进行ANOSIM 检验,结果显示两航次间小型底栖动物群落无显著差异(R=−0.07,p=0.741)。
3.2.3 小型底栖动物和环境因子相关性
对小型底栖动物丰度和沉积物有机质、重金属、黏土、粉砂、砂含量以及水深、温度和盐度等环境要素进行相关性分析,结果表明(表4):小型底栖动物总丰度和线虫丰度与重金属含量呈显著正相关(Cd除外);动吻类丰度和有机质含量以及Pb、Cr、Cu、Zn 含量呈极显著正相关,与As 和黏土含量呈显著正相关,与砂含量呈显著负相关;其他类群丰度和环境因子之间并无显著相关性。对各类群间丰度进行相关分析结果表明,小型底栖动物总丰度与线虫和动吻类丰度极显著相关,与桡足类丰度显著相关。相关分析结果表明,影响研究海域内小型底栖动物总丰度的主要类群为线虫、动吻类和桡足类,其中动吻类和沉积物中重金属含量密切相关,沉积物黏土和砂含量影响动吻类的丰度,线虫丰度与沉积物间重金属含量有较好的相关性。

图 6 研究海域小型底栖动物群落聚类分析Fig. 6 CLUSTER analysis of meiofaunal assemblages of the study area
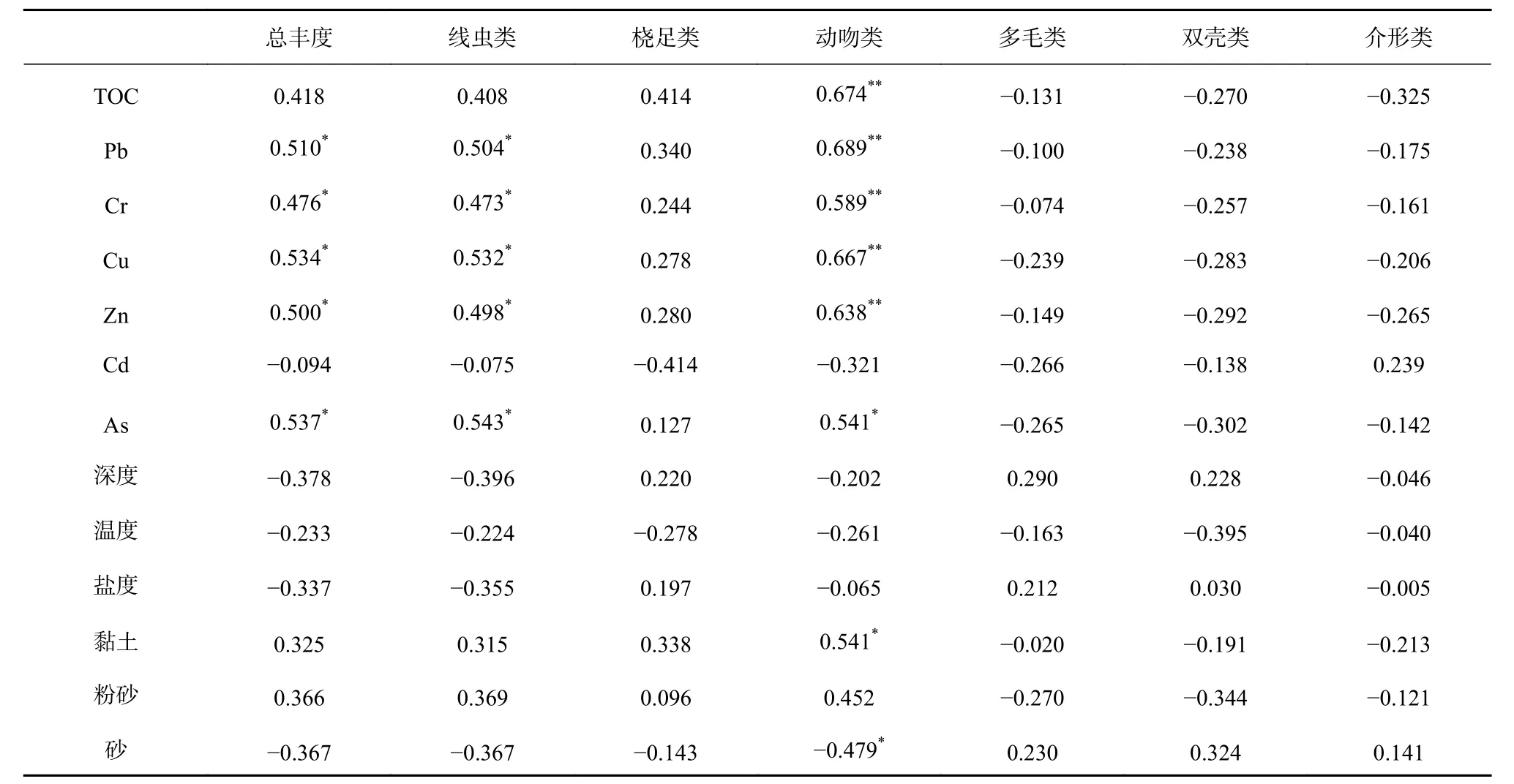
表 4 小型底栖动物丰度和环境因子相关性Table 4 Relative coefficient between meiofaunal abundance and environmental variables
对环境因子和小型底栖动物丰度进行BIOENV分析,结果表明:9 月航次影响小型底栖动物群落差异的最主要因素为水深和有机质组合,相关系数为0.329;12 月航次为有机质含量、砂含量和Zn 含量的组合,相关系数为0.774,除水深、温度、盐度及Cd 含量,其他因子组合也可以较好的解释站位间群落差异;从整体上,黏土含量和Pb、Cu、Zn 含量能够最好地解释研究海域站位间小型底栖动物的差异,相关系数为0.484,而水深和有机质含量与重金属组合也可以较好解释差异原因,相关系数为0.483。
4 讨论
4.1 长江冲淡水、台湾暖流以及沿岸流对底栖环境的影响
9 月航次各站位水体的温、盐性质存在显著差异和层化现象,这与长江径流的输入、黑潮入侵以及上升流等密切相关。受长江冲淡水影响,长江口及附近海域海表层水为高温、低盐的水,底层水为低温高盐水,如D1、C5 和C6 站位。许多研究已证明[29–30],在长江口31°00′~32°00′N,122°20′~123°10′E 之间存在上升流区,该上升流会将底层高盐低氧水携带至表层,因此通过垂向混合作用,C1 和C3 站位水体未表现出层化现象。从近岸向外海,C 断面底层水温逐渐降低,并在C6 站位达到最低值,而在D 断面随深度增加温度与盐度均显著升高,这与C、D 两断面所受海流影响不同有关。在C 断面,C5 和C6 站位受到海流的影响,其底层水来自南海沿岸流冷水与长江口底层的混合水,因此底层水温低于近岸站位[31–32]。有关台湾暖流及黑潮入侵的研究表明[8,33],闽浙近岸50 m 等深处底层水主要来自台湾东北部黑潮下层水(120~250 m 层)对东海陆架的入侵,温度特征值为19℃,盐度特征为34.4,与D4–D8 站位温盐特征相似。
与长江口海域相比较,浙江海域在冬季的温盐特征较稳定和简单。在12 月航次中,各站位水体未表现出明显的层化现象,但近岸站位由于受长江冲淡水南下分支以及季风的影响[30],其表层温度略低于底层温度,底、表盐度均低于外海站位。有关台湾暖流底层水在冬季入侵位置,相关研究已经证明该暖水可以到达31°N 或更远位置[7]。在本研究中,A、B 断面外海站位的温、盐特征表明,在冬季,台湾暖流水可以入侵至该纬度海域,并且是以均一性水体入侵,分布在整个深度。
4.2 小型底栖动物群落分布特征
4.2.1 上升流区和低氧对小型底栖动物群落的影响
先前研究已证明,在长江口31.00°~32.00°N,122.33°~123.17°E(图7)之间存在上升流区。韦钦胜等[34]在比较之前研究后发现,夏季(8 月)长江口低氧中心区大致位于30.83°~33.05°N,122.5°~123.5°E(图7)之间,刘海霞等[35]指出在进入9 月之后低氧区会向长江口南侧偏移并逐渐消失。
9 月航次中,小型底栖动物丰度最高值出现在D1 站位,正处于上升流和低氧重叠区,小型底栖动物的高丰度值和上升流密切相关。一方面,上升流将底层营养盐,特别是磷酸盐,携带至表层,有利于浮游植物大量繁殖[36];相关研究也表明,在长江口外上升流区存在叶绿素含量峰值[37–38],因此充足的食物来源是D1 站位小型底栖动物丰度较高的原因之一。另一方面,浮游植物大量沉降于表层沉积物,其降解会导致底氧快速消耗进而形成低氧环境[34,39]。低氧的出现有利于特别是海洋线虫等对低氧具有较高耐性的小型底栖动物繁殖[40–41],同时会迫使对氧敏感的大型底栖动物发生迁移甚至死亡[42],减小了小型底栖动物被捕食压力[43],利于小型底栖动物生存。在全球范围内,溶氧低值带小型底栖动物均表现出较高的丰度值[44–46]。
Hua 等[47]对长江口低氧带的研究也表明在该区域小型底栖动物具有较高的丰度值[高值达(3 699±1 239)个/(10 cm2)],长江口北侧向南侧存在丰度值增大的现象,与本研究情况相似。许多研究表明,小型底栖动物丰度和沉积物叶绿素a 的含量呈正相关性[20,23,47],C1 站位丰度值低于D1 站位可能与C1 站位叶绿素含量偏低有关,此外,C1 站位强烈的上升流和潮混合作用不利于小型底栖动物在沉积物中的固着和繁殖。

图 7 黄海沿岸流、黑潮分支、上升流和低氧中心区和小型底栖动物丰度分布Fig. 7 Water currents, upwelling, center of low oxygen area,and distribution of meiofauna
不同的小型底栖动物类群对低氧的耐受性显著不同,相关研究均表明海洋线虫对低氧不敏感而桡足类对低氧反应反敏感[43–44,48],Grego 等[49]低氧现场模拟实验表明,在低氧发生后底栖桡足类的丰度和多样性均显著下降。Hua 等[47]在分析长江口低氧区线虫和桡足类与溶氧的相关性是发现,线虫和溶氧之间无显著相关性,而桡足类与溶氧呈显著正相关。在本研究中,线虫和桡足类对低氧的耐受性亦可通过线虫与桡足类的丰度比值(N/C)来佐证,在低氧区,C1 和D1 站位N/C 比值显著高于其他站位(D4 站位除外)。在D4 站位N/C 比值异常高,与相邻站位比较,其在环境组成上并无显著差异,据此推测D4 站位可能存在低氧现象。
在C 断面,小型底栖动物丰度由近岸向外海显著增加,但线虫的优势度下降,小型底栖动物类群增多,特别是在C6 站位,小型底栖动物丰度最高,种类最多,这可能与C6 站位溶氧含量增加有关。同时,也在一定程度上反映出氧气可能是限制小型底栖动物群落物种多样性的关键因素。此外,C6 站位小型底栖动物的高丰度值和多样性可能与黄海冷水团的存在有关。与长江口近岸区海域相比,冷水团内部环境更加稳定,更有利于小型底栖动物的繁殖[50]。
4.2.2 黑潮对小型底栖动物群落的影响
相关研究表明[8]黑潮对东海陆架底层主要通过黑潮近岸分支(沿60 m 等深线)和黑潮离岸分支(沿100 m 等深线)两个分支入侵。通过分析各站位底、表层海水的温、盐特征可以发现,在9 月航次和12 月航次,除C 断面外,水深超过60 m 的站位底层温、盐特征均表现出黑潮水特征。对小型底栖动物的水平分规律进行比较可以发现,各断面小型底栖动物的丰度最高值均出现在水深60 m 附近(D1 除外),与黑潮近岸分支入侵路线高度一致(图7)。张志南等[51]对黄、东海小型底栖动物的研究中亦发现类似规律,小型底栖动物丰度在50 m 等深线处具有极大值,特别是在长江口以南东海陆架海域。
小型底栖动物丰度在60 m 等深线附近出现最高值可能是黑潮入侵的结果。食物来源是限制小型底栖动物丰度的重要因素之一。黑潮入侵为东海底层水带来丰富的磷酸盐和清澈的大洋水,在提供营养盐的同时改善了光照条件,有利于浮游植物生长繁殖和提高光合作用效率[52],使得该海域相对近岸站位具有更高的初级生产力,为小型底栖动物繁殖提供足够食物来源。同时黑潮水富含的溶氧也在一定程度上补充了东海底层水体氧的消耗,缓解了低氧现象的发生[53],有利于其他对底氧敏感的小型底栖动物类群的生存。因此,相对于近岸站位,小型底栖动物群落在该海域表现出高丰度值和多样性,如在N/C 比上,近岸站位(A1 和B1)N/C 比值均超过200,远高于黑潮入侵路线(A5 和B3)站位。此外,黑潮近岸分支的温度和盐度在季节间均具有较好的稳定性[54],为小型底栖动物提供了一个相对稳定的生存空间,可能也是造成其丰度较高的原因之一。
外海站位小型底栖动物丰度的下降可能是因为深度的增加,光照强度减弱,导致底栖藻类的生产力下降;同时随离岸距离的增加陆源营养物质输入减少可能进一步导致了生产力的下降。据此推测,食物来源可能是限制外海站位小型底栖动物丰度的重要因素。许多研究均发现,小型底栖动物丰度和沉积物叶绿素a 密切相关,叶绿素a 在一定程度上反映了小型底栖动物的食物来源的重要组成。目前,本研究中尚缺少沉积物叶绿素a 的数据,在后续调查中,补充沉积物中叶绿素a 的数据将有助有我们更加全面地认知黑潮入侵是如何对小型底栖动物群落的分布产生影响。
4.3 与同海域其他研究比较
与同海域其他研究比较,本研究在小型底栖动物类群组成、丰度以及水平分布上具有较好的延续性,但又存在少许差别(表5)。
在类群组成上,线虫均为最优势类群,其他依次为桡足类、动吻类、多毛类等;受低氧和近岸污染影响,在长江口近岸海域动吻类相对丰度较其他类群略高(同Hua 等[47]),而靠近外海站位桡足类和多毛类相对丰度略有增加。
在丰度组成上,本研究与华尔等[20]、Hua 等[47]、孟昭翠等[55]等研究在数值上最为接近,略高于其他,但显著高于张志南等[51]的研究。一方面,小型底栖动物的平均丰度受采样范围的影响,另一方面,不同的采样深度和网筛孔径影响小型底栖动物的获取效率。对比分析先前的研究可以发现,在长江口及其邻近海域,小型底栖动物的高丰度值均出现在30°~31°N,122.5°~123.5°E 之间,基本与长江口上升流区和低氧带重合。并且在长江口外,自北向南,自西向东部外海延伸至50~60 m 等深线处,小型底栖动物丰度表现出升高的趋势[23–24,47,56];同时,外海站位小型底栖动物丰度显著低于外海站位。在浙江以东的东海海域,小型底栖动物丰度最高值则出现在近岸站位[51]。因此,本研究中,小型底栖动物的平均丰度值较高可能与采样站位的分布有关。在垂直分布上,绝大多数小型底栖动物分布在沉积物的表层,张志南等[51]研究表明0~5 cm 的取样深度可以获取90%左右的小型底栖动物,而在渤海海域的取样效率甚至可达95%,因此更深的取样深度可能对小型底栖动物的获取效率提升并不显著。在本研究中,在浙江沿海和长江口等高丰度值站位,有大量个体细小的线虫出现(体长约280~300 μm),此类线虫在分选时易通过50 μm 孔径网筛但被31 μm 孔径网筛所截留,这可能是造成张志南等[51]研究小型底栖动物丰度显著低于其他研究的原因。

表 5 与同海域其他研究比较Table 5 Comparison with other studies in the same area
5 结论
(1)长江口外的上升流和低氧会导致该海域小型底栖动物丰度增加,多样性下降。上升流会导致该海域水体初级生产力增加,低氧会遏制大型底栖动物以及对低氧敏感的小型底栖动物类群(桡足类等)的生存,减小了小型底栖动物被捕食的压力,有利于海洋线虫的生存。
(2)黑潮入侵会导致小型底栖动物在其入侵路线上丰度和多样性增加。黑潮入侵的结果为小型底栖动物提供了充足的食物来源和溶氧,有利于小型底栖动物的繁殖和对溶氧敏感类群动物的生存。同时,黑潮水稳定的温、盐性质为小型底栖动物提供了稳定的生长环境,减少了季节变化所带来的不利影响。小型底栖动物在黑潮入侵路线上的高丰度值和多样性可视为其对黑潮入侵的响应。
(3)网筛孔径和采样深度以及采样范围是影响小型底栖动物平均丰度的重要因素。与同海域研究比较,不同航次间小型底栖动物的丰度差异主要受采样区域影响,与小型底栖动物在水平分布上呈现斑块状分布的特点有关。绝大多数小型底栖动物分布在沉积物表层和次表层;不同的网筛孔径对小型底栖动物的获取率差异明显,特别是对海洋线虫。
