九段沙湿地大型底栖动物分布特征及其影响因子
2020-03-12赵成建童春富
赵成建,童春富*
( 1. 华东师范大学 河口海岸学国家重点实验室,上海 200062;2. 崇明生态研究院,上海 200062)
1 引言
盐沼湿地广泛分布于中高纬度河口海岸地区,是世界上生产力最高的生态系统类型之一,具有重要的生态系统服务功能[1]。大型底栖动物是盐沼湿地中重要的生物类群,能为其他类群提供食物[2],还可以加速碎屑物的分解,调节沉积物与水界面的物质交换[3],是生态系统物质循环和能量流动的重要环节。
目前国内外关于盐沼湿地大型底栖动物已有较多的研究。早期的研究多集中于大型底栖动物群落的分布特征[4–5],之后也有不少学者对盐沼湿地大型底栖动物与环境因子之间的关系进行了探讨[6–7]。由于淹水频度、沉积物理化因子等随高程梯度变化而变化,高程梯度往往被认为是环境因子的综合表征[8–9]。有相当一部分研究已经针对大型底栖动物沿高程梯度的分布特征开展[10–12],但大部分工作集中在同一个区域,涉及不同区域的对比分析还比较少[11]。
九段沙为长江口的新生沙洲,有典型盐沼湿地发育。九段沙湿地大型底栖动物目前已经开展了部分研究,但主要集中于大型底栖动物群落的组成和时空分布特征等方面[13–15],而对大型底栖动物在不同区域、不同高程生境分布特征及其影响因子的研究较少,仅有少数学者对九段沙不同区域、不同高程生境大型底栖动物的分布特征进行了研究,而对其影响因子的研究并不完善[11]。因此,本文在已有研究的基础上,分析了九段沙不同沙体、不同高程生境大型底栖动物的群落组成特征,以及大型底栖动物与多种生境因子之间的关系,进一步揭示了大型底栖动物群落的空间分布特征及其影响因子。
2 研究区域概况与方法
2.1 研究区域概况
九段沙是长江口新生沙洲湿地,主要由江亚南沙、上沙、中沙与下沙4 部分组成,总面积为423.2 km2;该区域气候属亚热带季风性气候,年平均气温为15.7℃;1 月气温最低,月平均气温为3.3℃;7 月气温最高,月平均气温为27.4℃;年降水量为1 143 mm[16]。九段沙土著优势植物为芦苇(Phragmites australis)、海三棱藨草(Scirpus mariqueter)、藨草(Scirpus triqueter)。自1997 年引种互花米草(Spartina alterniflora)以来,互花米草分布范围持续扩张,分布面积已超过其他植物群落。九段沙底栖动物以软体动物和甲壳动物为主,主要优势种包括河蚬(Corbicula fluminea)、堇拟沼螺(Assiminea violacea)和无齿螳臂相手蟹(Chiromantes dehaani)等。
2.2 研究方法
2.2.1 样品采集
根据《海洋调查规范(GB/T 12763–2007)第6 部分:海洋生物调查》,在九段沙的江亚南沙、上沙和下沙设置4 条采样断面,每条断面沿高程梯度在不同植被带设置采样站点,于2016 年10 月进行取样调研。J1 断面位于江亚南沙,设置了光滩(J1B)、海三棱藨草带(J1Sm)和芦苇带(J1Pa)3 个采样站点;J2 断面位于上沙,同样设置了光滩(J2B)、海三棱藨草带(J2Sm)和芦苇带(J2Pa)3 个采样站点;J3 断面位于下沙南侧,设置了光滩(J3B)和互花米草(J3Sa)2 个采样站点;J4 断面位于下沙东南侧,设置了光滩(J4B)、海三棱藨草带(J4Sm)和互花米草(J4Sa)3 个采样站点,具体站点如图1 所示。
每个站点设置5 个与植被带平行的采样样地,每个样地设置2 个植被取样样方(50 cm×50 cm)、2 个大型底栖动物取样样方(25 cm×25 cm),每个样地采集2 个沉积物泥柱(30 cm)。取样时,用枝剪齐地剪取样方内植物,带回实验室于60℃烘干至恒重,称取干重,精确到0.01 g;现场记录植株密度、高度。在剪取了地上部分的植物样方内,设置25 cm×25 cm 大型底栖动物取样样方,先用镊子拣取表面的大型底栖动物,然后向下挖至30 cm 深,将挖出的泥样放入0.5 mm孔径的筛网内淘洗,将得到的底栖动物用5%的福尔马林溶液现场固定,带回实验室进行鉴定、计数,并在60℃烘干至恒重,称取干重,精确到0.000 1 g。另外在每个样方的邻近区域,用直径35 mm 的不锈钢采泥器采集1 个30 cm 长的沉积物柱状样,现场分割成6 段5 cm 长的小段,分别用自封袋保存后带回实验室,于60℃烘干至恒重,称重后计算容重。
2.2.2 土壤理化因子的测定
在采样的同时,采用氧化还原电位计(IQ 150)、土壤三参仪(WET-3)及盐度计现场测定记录沉积物的pH、氧化还原电位、温度、电导率和盐度等指标,每个指标在每个样地中重复测定2 次,沉积物含水率用干重和湿重之差除以湿重来计算。烘干后的沉积物剔除植物根系,用马尔文激光粒度仪(MS2000)测粒度,另外取一部分沉积物过100 目筛网,用元素分析仪(Vario EL III)测定总碳和总氮含量。
2.3 数据处理
采用相对多度(Relative Abundance,%N),相对生物量(Relative Biomass,%B)和Pinkas 相对重要性指数(Index of Relative Importance,IRI)来反映特定生境(植被带)大型底栖动物群落的组成特征[17]。

式中,ni,bi,fi分别为第i 种生物的个体数、生物量和出现次数;N、B、F 分别为总个体数、总生物量和总调研次数。
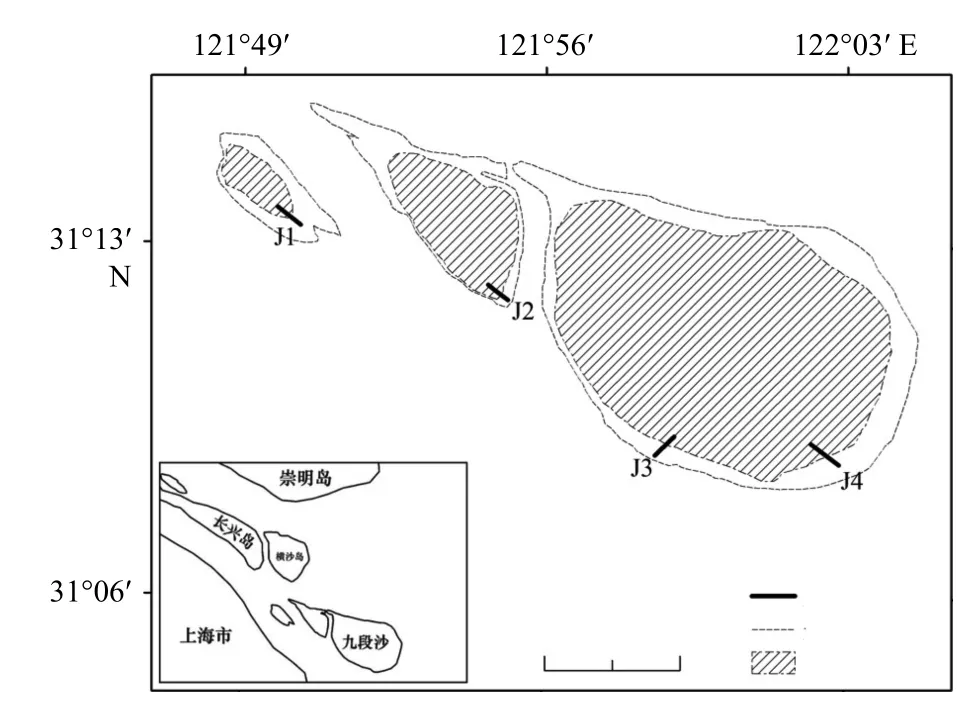
图 1 九段沙采样断面示意图Fig. 1 Map of sampling sections in Jiuduansha
数据采用统计软件包(IBM SPSS Statistics 19)进行处理。对数据先进行方差齐性检验,不具备方差齐性的数据需转化后再进行分析。采用单因素方差分析(one-way ANOVA)对同一断面不同高程生境和不同断面同一生境类型大型底栖动物的多度、生物量的差异进行显著性分析,再选用最小显著差法(LSD)进行均数间多重比较[18]。以P<0.05 表示显著差异,P<0.01 表示极显著差异。大型底栖动物多度、生物量与生境因子之间的关系采用统计软件PRIMER 5.2.8 进行非线性回归分析。用BVSTEP 程序找出与目标变量匹配最好的环境因子组合,并计算相关系数;再用RELATE 程序检验两者之间的相关性是否显著。所有数据在使用时需经过4 次方根变换,以满足数据的正态性要求;环境因子的相关数据需要进行标准化处理,以避免量纲差异造成的影响[19]。
用统计软件PRIMER5.2.8 对不同断面、不同高程生境(植被带)的大型底栖动物(物种−密度数据)进行聚类和非度量多维尺度分析(Non-metric Multi-dimensional Scaling, nMDS)。
3 研究结果
3.1 大型底栖动物群落组成
调研期间共采集到大型底栖动物14 种,隶属于3 门4 纲6 目12 科。其中,软体动物8 种,占总物种数的57.14%;环节动物和甲壳动物各3 种,分别占总物种数的21.43%。
不同断面、不同植被带优势种组成有一定差异,如表1 所示。J1 断面光滩和海三棱藨草带的主要优势种均为河蚬(IRI>20),而芦苇带的优势种为堇拟沼螺与红螯螳臂相手蟹(Chiromantes haematocheir)(IRI>20)。J2 断面光滩的优势种为河蚬与堇拟沼螺(IRI>20),海三棱藨草带的优势种为河蚬(IRI>20),芦苇带的优势种为中华拟蟹守螺(Cerithidea sinensis)(IRI>20)。J3 断面光滩的主要优势种为黑龙江河篮蛤(Potamocorbula amurensis)(IRI>20),堇拟沼螺也具有一定的优势特征(10<IRI<20);互花米草带的主要优势种为堇拟沼螺(IRI>20),无齿螳臂相手蟹也具有一定的优势特征(10<IRI<20)。J4 断面光滩和海三棱藨草带的主要优势种也为黑龙江河篮蛤(IRI>20),互花米草带的主要优势种为堇拟沼螺(IRI>20),无齿螳臂相手蟹也具有一定的优势特征(10<IRI<20)。
3.2 大型底栖动物的分布特征
大型底栖动物的密度如图2A 所示。J1、J4 断面中,不同高程生境大型底栖动物的密度存在显著差异(P<0.05),而J2、J3 断面中,不同高程生境大型底栖动物密度的差异均不显著(P>0.05)。两两比较的结果显示,J1 断面光滩和海三棱藨草带大型底栖动物的密度均显著高于芦苇带(P<0.05),但光滩和海三棱藨草带之间的差异不显著(P>0.05);J4 断面海三棱藨草带大型底栖动物的密度显著高于互花米草带和光滩区域(P<0.05),而互花米草带大型底栖动物的密度也显著高于光滩区域(P<0.05)。
大型底栖动物的生物量如图2B 所示。除了J3 断面,其他3 个断面中,不同高程生境大型底栖动物的生物量均存在显著差异(P<0.05)。两两对比的结果显示,J1 断面的光滩和海三棱藨草带大型底栖动物生物量的差异不显著(P>0.05),但光滩和海三棱藨草带的生物量均显著高于芦苇带(P<0.05);J2 断面光滩和海三棱藨草带大型底栖动物生物量的差异也不显著(P>0.05),但芦苇带的生物量显著高于光滩和海三棱藨草带(P<0.05);J4 断面海三棱藨草带大型底栖动物的生物量显著高于光滩和互花米草带(P<0.05),但光滩和互花米草带之间的差异不显著(P>0.05)。
不同断面相同生境类型大型底栖动物的密度均存在显著差异(P<0.05)。两两比较的结果显示,在光滩生境中,J3 断面大型底栖动物的密度显著高于J1、J2 和J4 断面(P<0.05),但J1、J2、J4 断面之间的差异不显著(P>0.05);在海三棱藨草生境中,J4 断面大型底栖动物的密度显著高于J1 和J2 断面(P<0.05),但J1 和J2 断面之间的差异不显著(P>0.05);在芦苇/互花米草生境中,分布有互花米草的J3 和J4 断面大型底栖动物的密度显著高于分布有芦苇的J1 和J2 断面(P<0.05),但J1 和J2 断面之间、J3 和J4 断面之间差异均不显著(P>0.05)。
从大型底栖动物的生物量来看,不同断面相同生境类型间也都存在显著差异(P<0.05)。两两比较的结果显示,在光滩区域,J1 断面大型底栖动物的生物量显著高于其他3 个断面(P<0.05),但J2、J3、J4 这3 个断面之间都不存在显著差异(P>0.05);在海三棱藨草带,J1 和J4 断面大型底栖动物的生物量没有显著差异(P>0.05),但都显著高于J2 断面(P<0.05);在芦苇/互花米草带,J2 断面大型底栖动物的生物量显著高于其他3 个断面(P<0.05),但J1、J3、J4 断面两两之间的差异都不显著(P>0.05)。
不同站点大型底栖动物群落组成的聚类和非度量多维尺度分析的结果如图3 所示。聚类分析将11 个站点分成了3 组,J2Pa 站点单独成为一组;光滩和海三棱藨草带的7 个站点组成一组;高潮位植被带的3 个站点组成一组,其中,J3Sa 和J4Sa 这2 个互花米草带的大型底栖动物群落相似性达到了80% 以上。nMDS 分析图的Stress 值为0.06,说明解释站点之间相似关系的可信度较为一般,但仍可以把11 个站点大致分成3 组,这与聚类分析的结果较为一致。聚类和非度量多维尺度分析结果表明,与高程对应的植被带是表征大型底栖动物群落组成差异的主要生境特征。
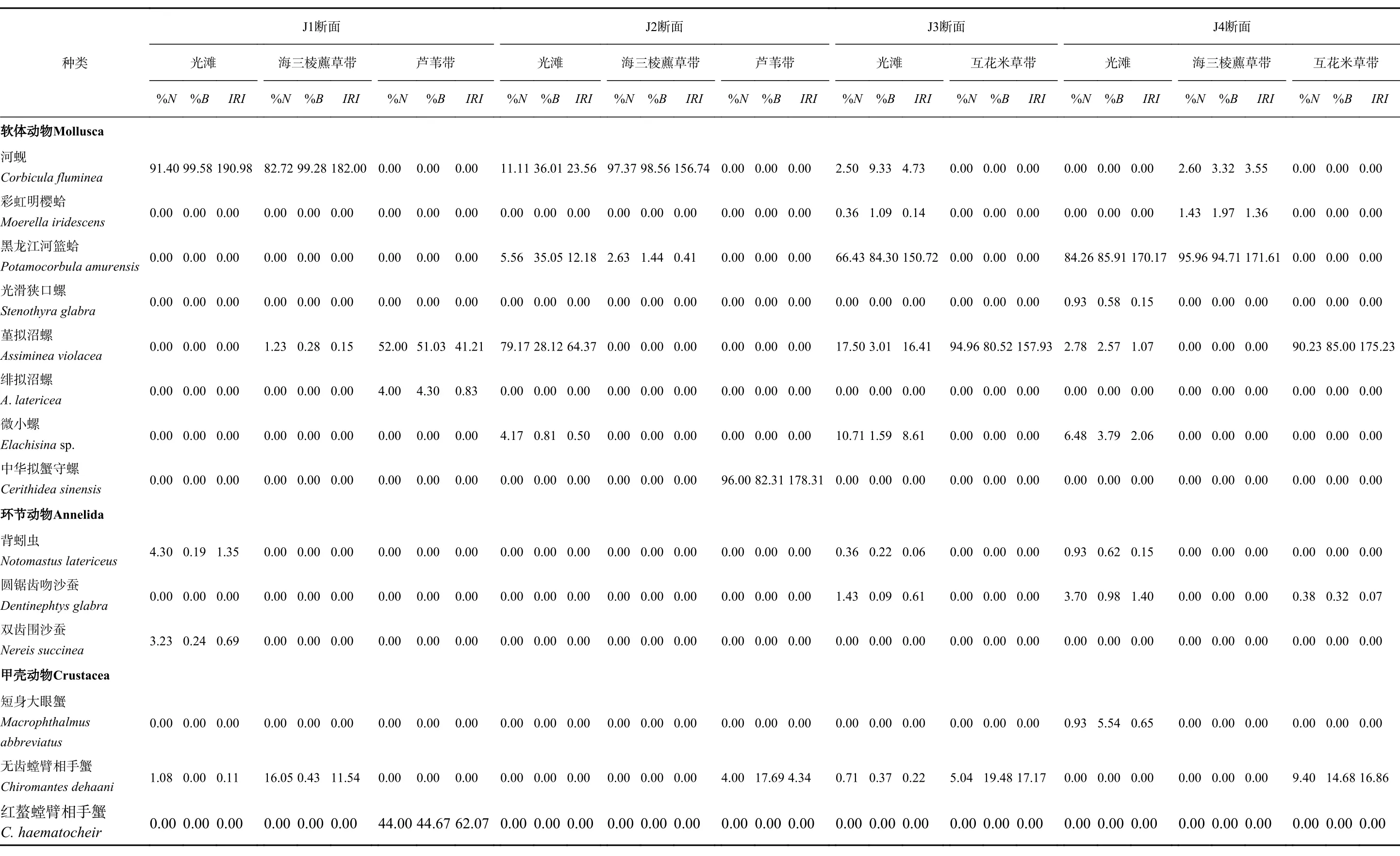
表 1 九段沙大型底栖动物群落组成特征Table 1 Community composition of the benthic macroin vertebrates in Jiuduansha
3.3 不同站点主要生境特征
不同站点的植被特征如表2 所示。海三棱藨草仅植株密度在不同站点间存在显著差异(P<0.05),其中J1、J2 断面与J4 断面之间的差异均显著(P<0.05)。芦苇仅植株密度在J1 和J2 断面间存在显著差异(P<0.05);互花米草仅地上生物量在J3 和J4 断面间存在显著差异(P<0.05)。
不同站点的沉积物理化指标如图4 所示。J1 断面光滩和海三棱藨草带的沉积物盐度、pH、氧化还原电位与芦苇带之间的差异均显著(P<0.05);J2 断面的沉积物盐度在3 个生境间的差异均显著(P<0.05),光滩和海三棱藨草带的沉积物pH、氧化还原电位及电导率与芦苇带之间的差异均显著(P<0.05);J4 断面的沉积物氧化还原电位在3 个生境间的差异均显著(P<0.05),光滩和海三棱藨草带的沉积物盐度与互花米草带之间的差异均显著(P<0.05);除个别情况外,各断面不同生境同一深度的沉积物中值粒径之间的差异均显著(P<0.05)。
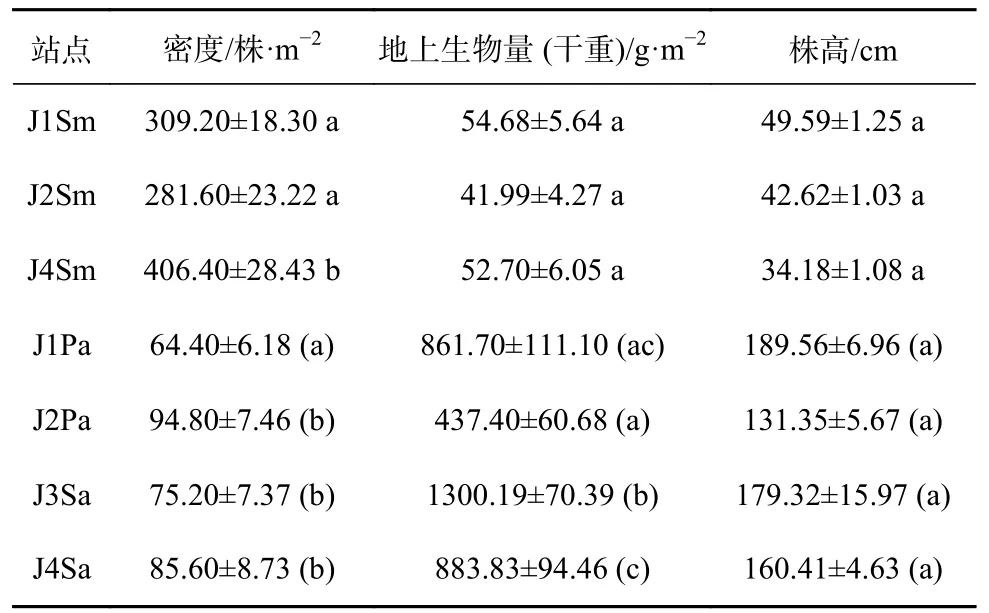
表 2 不同站点植株密度、生物量和高度(平均值±标准误)Table 2 Density, biomass and height of the plants in the different sampling sites(mean ± SE)
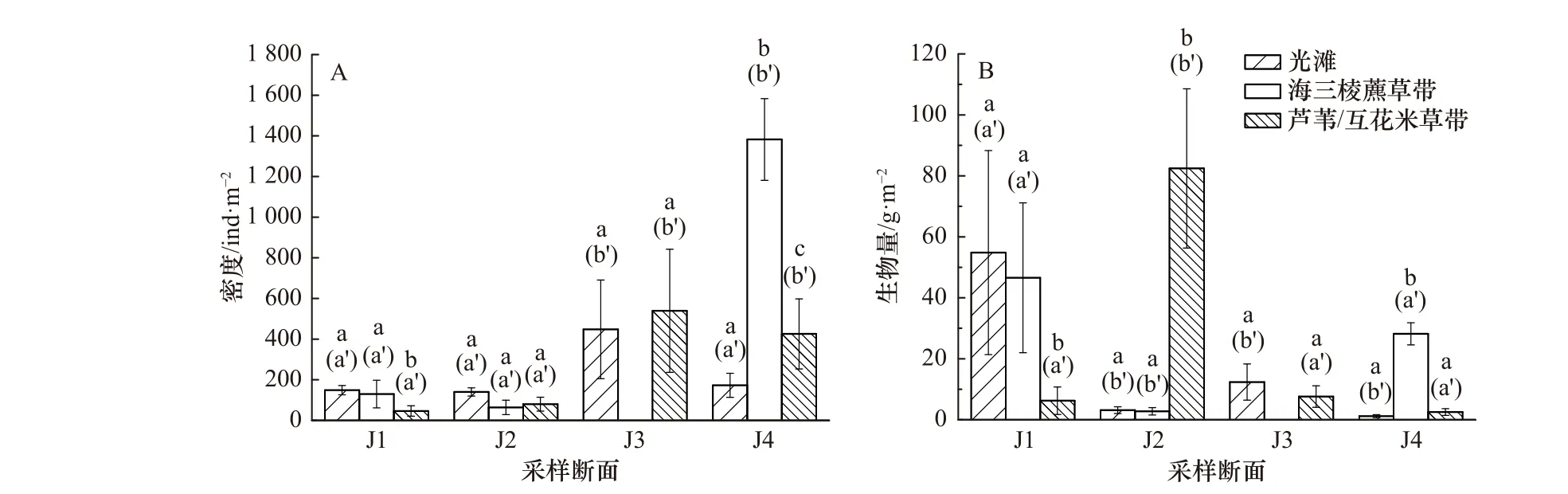
图 2 大型底栖动物的密度(A)和生物量(B)(平均值±标准误)Fig. 2 Density (A) and biomass (B) of the benthic macroinvertebrates (mean±SE)

图 3 大型底栖动物群落的聚类分析(a)及非度量多维尺度分析(b)Fig. 3 Cluster (a) and nMDS analysis (b) of the benthic macroinvertebrate community
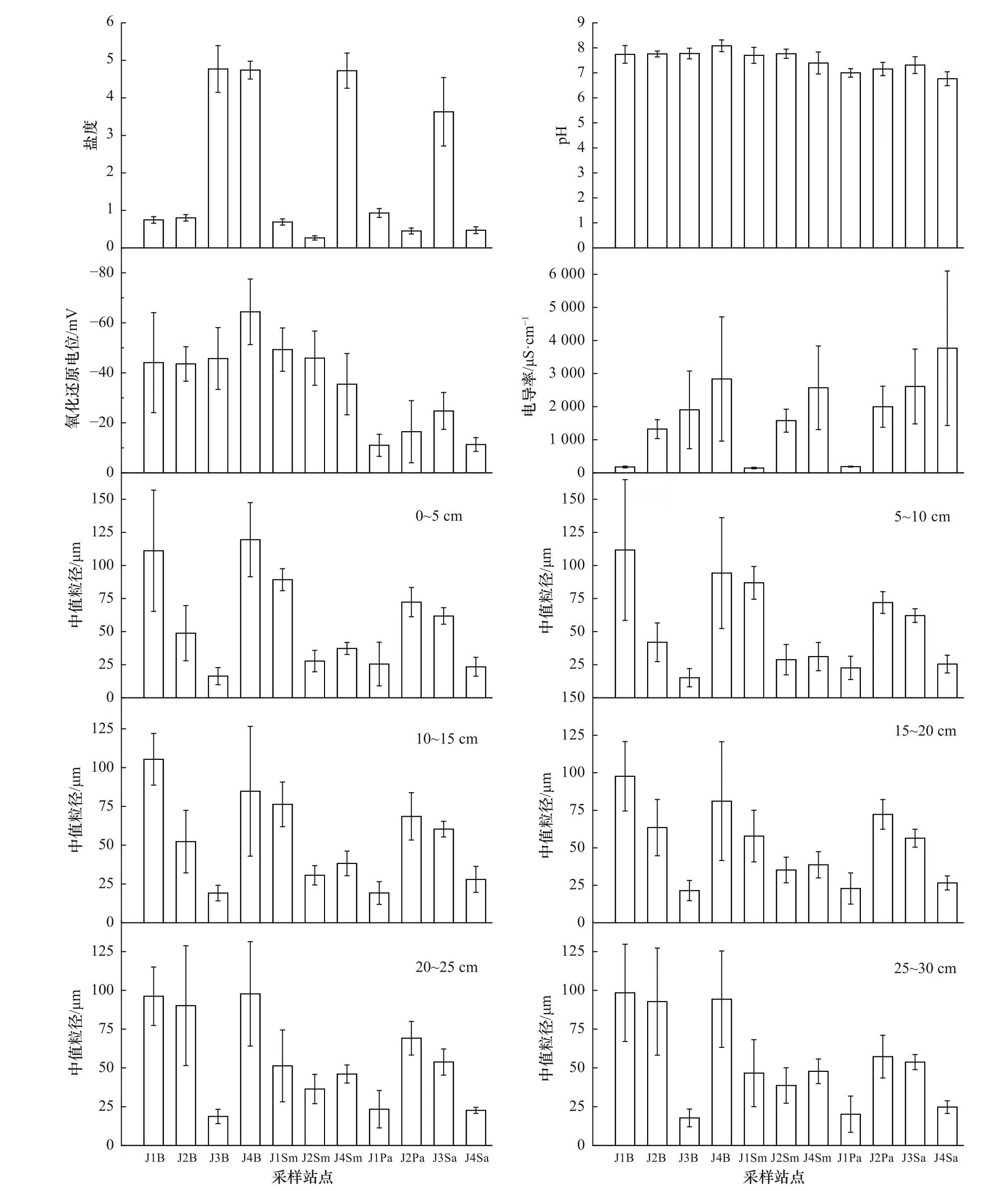
图 4 沉积物理化特征(平均值±标准误)Fig. 4 Physical and chemical characteristics of the sediments (mean±SE)
主要的沉积物理化指标在不同断面的相同生境类型间也有差异。在光滩区域,J3 和J4 断面的沉积物盐度显著高于J1 和J2 断面(P<0.05),J4 断面的沉积物pH 和氧化还原电位显著高于其他3 个断面(P<0.05),J4 断面的沉积物电导率显著高于J1 和J2 断面(P<0.05),除个别情况外,同一深度的沉积物中值粒径在不同断面间的差异均显著(P<0.05);在海三棱藨草带,沉积物盐度和电导率在三个断面间的差异均显著(P<0.05),J1 断面的沉积物氧化还原电位与J4 断面的差异显著(P<0.05),除15~20 cm 的沉积物中值粒径外,其他深度的沉积物中值粒径在不同断面间的差异均显著(P<0.05);在芦苇/互花米草带,J3 断面的沉积物盐度和pH 显著高于其他3 个断面(P<0.05),J2 和J3 断面的沉积物氧化还原电位显著高于J1 和J4 断面(P<0.05),J4 断面的沉积物电导率显著高于其他3 个断面(P<0.05),而J2 和J3 断面的沉积物电导率又显著高于J1 断面(P<0.05),除0~5 cm 的沉积物中值粒径外,其他深度的沉积物中值粒径在不同断面间的差异均显著(P<0.05)。
3.4 大型底栖动物与生境因子之间的关系
大型底栖动物的多度、生物量与生境因子之间的非线性回归分析结果如表3 所示。仅在现有断面的个别生境,部分生境因子的组合与大型底栖动物的多度或生物量表现出显著的非线性相关,且不同断面、相同生境的生境因子组合也有所不同。
4 讨论
4.1 不同高程生境大型底栖动物的分布
已有研究表明,河口潮间带大型底栖动物沿高程分布具有一定的规律性:在低潮带,由于淹水时间长,栖息在表层且运动能力较弱的物种难以生存,而栖息在沉积物深处的环节动物适宜在此生存[12];在中潮带,海三棱藨草在一定程度上改造了生境条件,一方面为底栖动物提供了生境的复杂性[20],而且海三棱藨草的地下根、茎也可以为底栖动物提供丰富的食物来源[21],另一方面,海三棱藨草通过减弱波浪的作用使运输的沉积物变细[22],从而使生境更加稳定,更适于软体动物的生存;在高潮带,淹水时间较短,沉积物稳定性强,适合掘穴洞居型的蟹类生存[23]。在本研究中,4 个断面低潮带环节动物的密度均高于中潮带和高潮带,中潮带软体动物的物种数均高于低潮带和高潮带,高潮带蟹类的密度和生物量均高于低潮带和中潮带,这些结果基本都与前人的研究一致[12]。
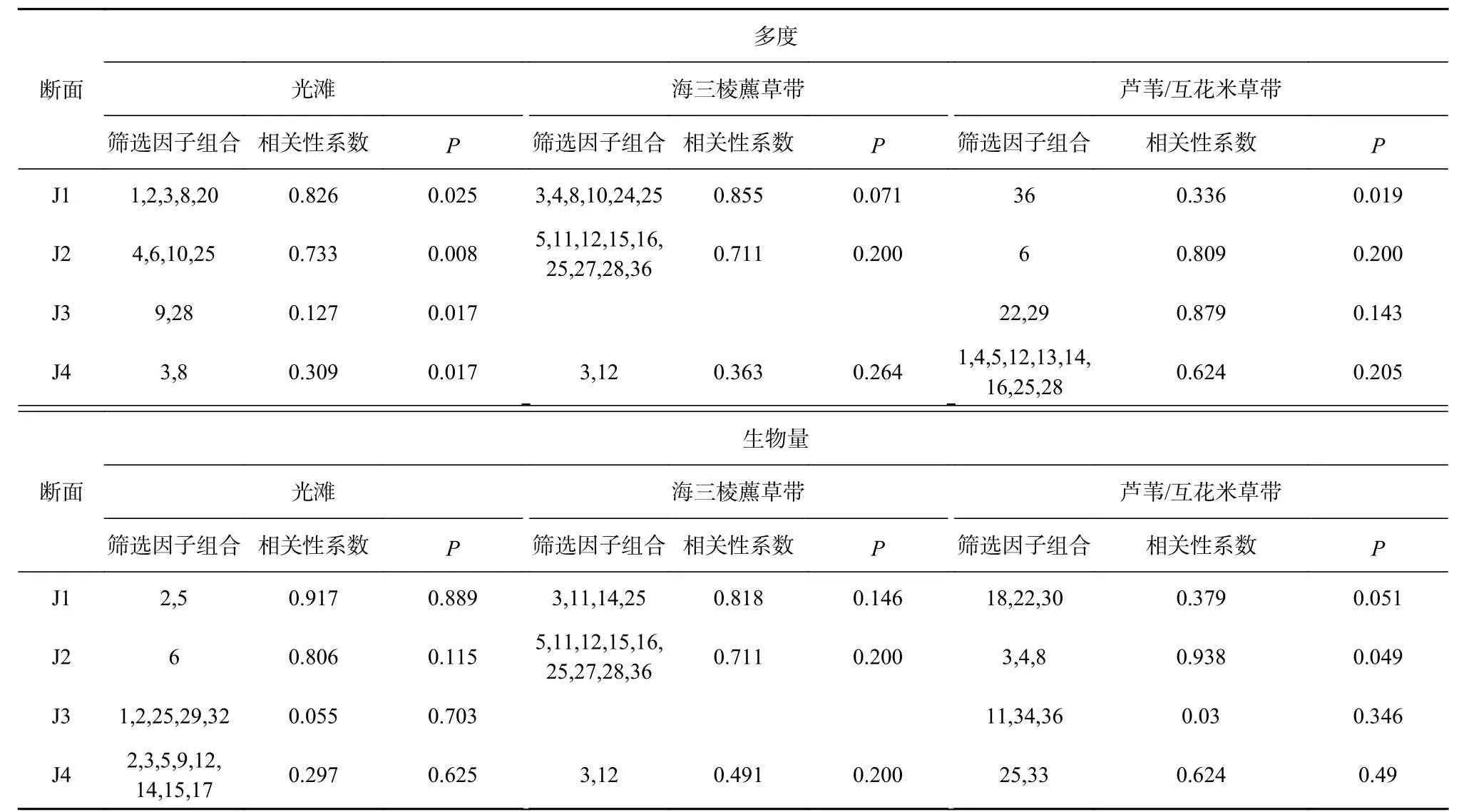
表 3 大型底栖动物与生境因子的非线性回归分析Table 3 Nonlinear regression analysis among the benthic macroinvertebrates and habitat factors
现有九段沙大型底栖动物空间分布特征的研究结果有很大差异,大部分研究都认为中潮带大型底栖动物的密度最高[23–25],但有研究发现高潮带大型底栖动物的生物量最高[24],也有研究证明低潮带大型底栖动物的生物量最高[25],还有研究发现中潮带大型底栖动物的密度和生物量均最高[23]。在本研究中,不同区域断面的情况存在明显差异。J1 断面低潮带大型底栖动物的密度、生物量均高于中潮带和高潮带;J2 断面大型底栖动物密度最高的是中潮带,而生物量最高的则是高潮带;J4 断面中潮带大型底栖动物的密度、生物量均高于低潮带和高潮带。之前的研究大部分都集中在同一个区域,生境条件的差异较小,而本研究涉及多个区域,区域之间的生境条件差异较大,J1 断面低潮带沉积物孔隙水的平均盐度为0.7,中值粒径为103 μm,显著大于中潮带和高潮带(P<0.05),低盐度和相对较大的沉积物中值粒径等条件非常适于河蚬的生存,大量河蚬在此栖息,因此J1 断面低潮带光滩大型底栖动物的密度和生物量均高于中潮带和高潮带;J2 断面高潮带大型底栖动物的生物量显著高于其他断面的所有生境(P<0.05),可能是由于上沙成陆时间较早,中华拟蟹守螺分布数量较多;J4 断面中潮带沉积物孔隙水的盐度较高,平均盐度为4.7,显著高于其他断面(P<0.05),较高的盐度适于黑龙江河篮蛤的生存,另外中潮带海三棱藨草的植株密度也显著高于其他断面(P<0.05),提供了更稳定的生境和更多的食物,滋生了大量的软体动物,因此中潮带大型底栖动物的密度和生物量均高于低潮带和高潮带。基于上述分析可知,不同区域由于环境条件的差异,大型底栖动物沿高程梯度的分布特征也存在明显差异。
4.2 生境因子与大型底栖动物分布
盐沼湿地底栖动物的分布受到多种因子的共同影响,包括沉积物盐度、中值粒径和植被类型等[26–28]。在本研究中,对大型底栖动物的多度和生物量有显著影响的生境因子主要是沉积物的中值粒径、氧化还原电位、总氮含量以及植株密度等。已有研究表明,大型底栖动物的分布受到沉积物粒径的重要影响[29],在本研究中,不同站点沉积物中值粒径的差异显著(P<0.05),沉积物粒径分布的差异对大型底栖动物的分布产生了显著影响。盐度一般被认为是影响大型底栖动物的一个重要因子[30],但在本研究中,盐度对大型底栖动物多度和生物量的影响并不显著,这可能是由于九段沙处于长江口咸淡水交汇处,底栖动物具有广盐性这一特征[31]。对于九段沙底栖动物与生境因子之间的关系,之前也有学者进行了研究,发现九段沙底栖动物的密度受到沉积物含水率的显著影响,但生物量与生境因子之间的相关性并不高[25],本文的研究结果与之类似,包括沉积物含水率在内的多种生境因子对多个生境大型底栖动物的多度产生了显著影响(P<0.05),但仅有J2 断面芦苇带大型底栖动物的生物量受到沉积物中值粒径等因子的显著影响(P<0.05)。另外,本研究还发现互花米草带大型底栖动物的密度显著高于芦苇带(P<0.05),可能是由于互花米草的单位面积地上生物量比芦苇高,能为大型底栖动物提供更多的食物,有利于大型底栖动物的生存。有研究发现,互花米草的入侵会改变沉积物的理化性质,如温度、盐度和中值粒径等,从而影响大型底栖动物的分布特征[32],本研究也发现,同一断面互花米草带沉积物的氧化还原电位、孔隙水盐度、中值粒径等理化因子与光滩和海三棱藨草带均存在显著差异(P<0.05),这些理化因子的差异导致了大型底栖动物分布特征的差异,这与前人的研究结果基本一致。在本研究中,不同断面、不同生境中对大型底栖动物的分布特征有显著影响的生境因子并不完全相同,原因是这4 个断面所在的位置不同,生境因子之间存在一定的差异。
综上所述,大型底栖动物沿高程梯度的分布具有一定规律性:低潮带环节动物的多度和生物量均高于中潮带和高潮带,中潮带软体动物的物种数均高于低潮带和高潮带,高潮带甲壳动物的多度和生物量均高于低潮带和中潮带,这也反应了生境因子沿高程梯度的变化。不同区域由于环境条件的差异,大型底栖动物沿高程梯度的分布特征也存在明显差异,对大型底栖动物的分布特征有显著影响的生境因子也有所差异。在今后的研究中,需要考虑到不同区域、不同高程之间环境条件的差异,进一步探讨大型底栖动物沿高程梯度的分布特征以及生境因子对大型底栖动物分布的综合影响。
