菜心耐热性评价及酶促抗氧化系统对高温胁迫的响应
2020-03-07庞强强孙晓东蔡兴来
庞强强,周 曼,孙晓东,张 文,蔡兴来
(海南省农业科学院 蔬菜研究所,海南省蔬菜生物学重点实验室,海南省瓜菜育种工程技术研究中心,海南 海口571100)
随着全球平均气温的升高,高温已成为影响农作物生产最重要的气候因子之一[1]。菜心(BrassicaparachinensisL.),又名菜薹,为十字花科芸薹属一、二年生草本植物,由白菜亚种长期选择和栽培驯化而来,是我国华南地区的特产[2]。菜心性喜凉,生长适温15~25 ℃,在30 ℃以上的高温天气下,菜心的生长发育就会受到影响,会导致菜苔细小、质劣,单株质量和商品产量大幅下降[3]。由于海南特殊的地理位置,季节差异大,温光水热资源分布不均,造成了夏秋高温多雨、冬春干旱少雨的不利气候条件,且全年高温期长,夏季田间温度可达40 ℃以上,高温已成为限制该地区菜心正常生长的重要因素。筛选和培育耐热性强的菜心种质是解决上述问题的必由之路,而耐热性鉴定是选育耐热品种的重要手段。已有研究表明,田间鉴定采用热害指数可有效区分菜心的耐热性,且其结果与室内鉴定基本一致[4-5]。
植物对于高温胁迫的响应并非完全是被动的,在长期的进化过程中,植物能够迅速感知外界温度变化并做出响应,以降低不利环境所造成的危害,维持体内基本的代谢反应[6]。例如,通过细胞膜稳定性、酶抗氧化系统、非酶活性氧清除剂等的变化以获得耐热性,从而减缓高温对自身的伤害。目前,高温胁迫生理研究在芸薹属作物如大白菜(Brassicarapassp. pekinensis)[7]、甘蓝(BrassicaoleraceaL.)[8]、花椰菜(BrassicaoleraceaL.)[9]、油菜(BrassicaoleraceaL.)[10]等广泛开展。相对而言,国内外对菜心耐热性的研究报道还较少。为此,本研究以19份菜心品种为试材,以热害指数为评价指标,对其进行田间耐热性鉴定。在此基础上,研究高温胁迫对耐热性不同材料叶片的电导率、丙二醛含量、抗氧化酶活性和相关基因表达的影响及胁迫解除后它们的恢复能力,旨在为菜心耐热品种的选育及抗逆栽培提供参考依据。
1 材料与方法
1.1 田间试验材料及试验设计
本试验以收集的19份菜心品种为试验材料,并对其进行编号CX1~CX19,来源见表1。田间试验于2017年7—8月在海南省农业科学院蔬菜研究所永发试验基地进行,室内试验在海南省蔬菜学重点实验室进行。以珍珠岩为基质进行穴盘育苗,当幼苗长至3叶1心时,选取生长一致的健壮幼苗移栽到田间,记录田间气温变化,其他管理均按常规。田间移栽后7和14 d选取上述材料15株进行热害症状调查。
1.2 人工模拟高温胁迫试验材料及试验设计
根据菜心田间耐热性筛选结果,选取耐热型品种CX10、中等耐热型品种CX17和不耐热型品种CX13用于人工模拟高温胁迫试验材料。选择饱满、均匀、无病虫害的种子播种在16孔穴盘内进行育苗,每穴播3粒种子,每个品种播12盘,共36盘,播种后采用常规栽培模式管理,长出1片真叶时进行间苗,每孔留苗1株。当幼苗长至3叶1心时,移入25 ℃/18 ℃(昼/夜),12 h/12 h(昼/夜),光照80%,湿度70%的人工气候箱(宁波海曙赛福实验仪器厂)中预培养3 d,之后将其中27盘(每个品种各9盘)幼苗移入另一人工气候箱内进行高温胁迫处理,处理温度为37 ℃/27 ℃(昼/夜),高温处理5 d后进行恢复处理。另外9盘(每个品种各3盘)幼苗仍然培养在25 ℃/18 ℃(昼/夜)的人工气候箱内(对照)。除温度不同外,高温胁迫处理的其他环境条件均与预培养时保持一致。分别取高温处理0 d(CK,即预培养条件下正常生长的植株)、3 d(H3)、5 d(H5)及恢复处理3 d(R3)菜心相同部位叶片,每个处理随机选取10株幼苗用于各项生长生理指标测定和RNA提取,试验均设3次重复。

表1 菜心品种名称及来源Table 1 Variety name and source of Brassica parachinensis L.
1.3 试验方法
1.3.1 热害指数调查
参照张景云等[11]、李光光等[4]的方法,略有改动,热害分级标准为:0级(无热害症状,植株生长正常);1级(植株受害叶片数<全株叶片数<1/3);3级(全株叶片数的1/3≤植株受害叶片数≤全株叶片数的1/2);5级(全株叶片数的1/2≤植株受害叶片数≤全株叶片数的2/3);7级(植株受害叶片数≥全株叶片数的2/3);9级(植株失去经济价值或死亡)。热害指数=∑(X×Xi)/(A×N),X代表热害级数,Xi代表X热害级数下的植株数,A代表最高级数,N代表调查总株数。
1.3.2 田间温度统计
采用温湿度自动记录仪(浙江托普云农科技股份有限公司,TPJ-20-LG)记录菜心生育期间田间气温变化,统计每天最高和最低气温。
1.3.3 生理指标测定
电导率、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性和过氧化氢酶(CAT)活性参照李合生等[12]方法。
1.3.4 基因表达分析
基因表达分析采用TransZol Plant试剂盒提取样品的总RNA,利用PrimeScriptTMRT reagent Kit with gDNA Eraser (Perfect Real Time)试剂盒反转录合成cDNA第一链。实时荧光定量反应程序按宝生物公司的SYBR®Premix Ex TaqTM (Tli RNaseH Plus)试剂盒说明书要求进行,以Actin为内参基因,目的基因的表达量用2-△△Ct法来计算,每个基因的表达反应重复3次。Mn-SOD、CAT和POD基因的引物序列根据与菜心种属较近的同源序列比对设计,引物由上海生工生物工程有限公司合成,序列信息见表2。
1.4 数据统计及分析
采用Excel软件计算每个材料各处理测定值与对照测定值的相对值,当相对值大于1时代表处理测定值高于对照测定值,相对值小于1时代表处理测定值低于对照测定值,相对值等于1时代表处理测定值等于对照测定值,之后再对数据进行作图。采用SPSS22.0对数据进行统计分析,单因素分析采用Duncan法。

表2 引物序列信息Table 2 Primer sequence information
2 结果与分析
2.1 菜心生育期间田间气温变化
由图1可知,菜心生育期间田间每天的最低气温在26~30 ℃,其中最低气温在28 ℃及以上的天数有57 d(91.94%)。最高气温在37 ℃及以上的天数有51 d(73.91%),40 ℃以上的天数有15 d(24.19%),最高气温可达41 ℃,说明菜心在整个生育期受到持续高温胁迫。
2.2 菜心田间热害指数调查结果
菜心田间热害指数调查结果见表3,通过2次热害指数的调查,计算平均值,依据热害指数平均值筛选出耐热性强的品种,指数越小说明材料耐热性越强。由表3可知,参试菜心各品种在田间自然高温条件下均出现热害症状,但不同品种间存在差异。耐热性由强到弱依次为CX9>CX3=CX19>CX10>CX14>CX6>CX11=CX16>CX5>CX1>CX17=CX18>CX12=CX15>CX4=CX8>CX7>CX13>CX2。利用筛选出的耐热材料和不耐热材料用于后续生理指标的测定。
2.3 聚类分析
根据热害指数,采用中间距离法对19个菜心品种进行聚类分析。由图2可知,19个菜心品种划分为耐热型、中等耐热型和不耐热型3种类型,其中耐热型品种有5个,分别为CX3、CX19、CX10、CX14、CX9;中等耐热型品种有11个,分别为CX11、CX16、CX6、CX17、CX18、CX1、CX5、CX12、CX15、CX4、CX8;不耐热型品种有3个,分别为CX7、CX13、CX2。
2.4 高温胁迫及恢复对菜心电导率和丙二醛含量的影响

图1 菜心生育期间田间最高气温和最低气温变化Fig.1 The highest and lowest temperature changes during the growth period of Brassica parachinensis L.

表3 热害指数调查结果Table 3 Investigation results of heat injury index

图2 十九个菜心品种热害指数聚类分析Fig.2 The clustering analysis of heat injury index of 19 cultivars
由图3可知,高温处理3 d时,CX10、CX17、CX13这3个菜心品种的电导率分别是对照的1.31、2.04、2.68倍,MDA含量分别是对照的2.32、3.68、4.03倍,其中耐热型品种CX10的电导率和MDA含量较对照上升幅度较小,其次为中等耐热型品种CX17,不耐热型品种CX13的上升幅度最大。高温处理5 d时,3个菜心品种的电导率和MDA含量仍显著高于对照。恢复处理3 d后,电导率和MDA含量较高温胁迫时有下降趋势,但仍高于对照处理,其中耐热型品种的变化幅度较小,不耐热型品种的变化幅度较大。由此可见,耐热型品种在高温胁迫下受到的伤害较小,并且恢复能力也远远强于不耐热型品种。
2.5 高温胁迫及恢复对菜心抗氧化酶活性的影响
由表4可见,在高温处理下,3个菜心品种叶片中SOD、POD、CAT活性变化一致,均呈先上升后下降的趋势。在高温处理3 d时,耐热型品种CX10的SOD、POD、CAT活性分别是对照的4.22、2.56和1.84倍,中等耐热型品种CX17的SOD、POD、CAT活性分别是对照的3.17、2.21和1.62倍,不耐热型品种CX13的SOD、CAT和POD活性分别是对照的2.35、1.30和1.27倍。在整个高温处理期间,无论指标的变化趋势如何,不耐热型品种CX13的抗氧化酶活性始终低于耐热型品种CX10,且前者的增加幅度均小于后者。在胁迫恢复3 d时,以上各指标均下降,但仍高于对照,其中各抗氧化酶活性均在耐热型品种中最高,其次为中等耐热型品种,在不耐热型品种中最低。
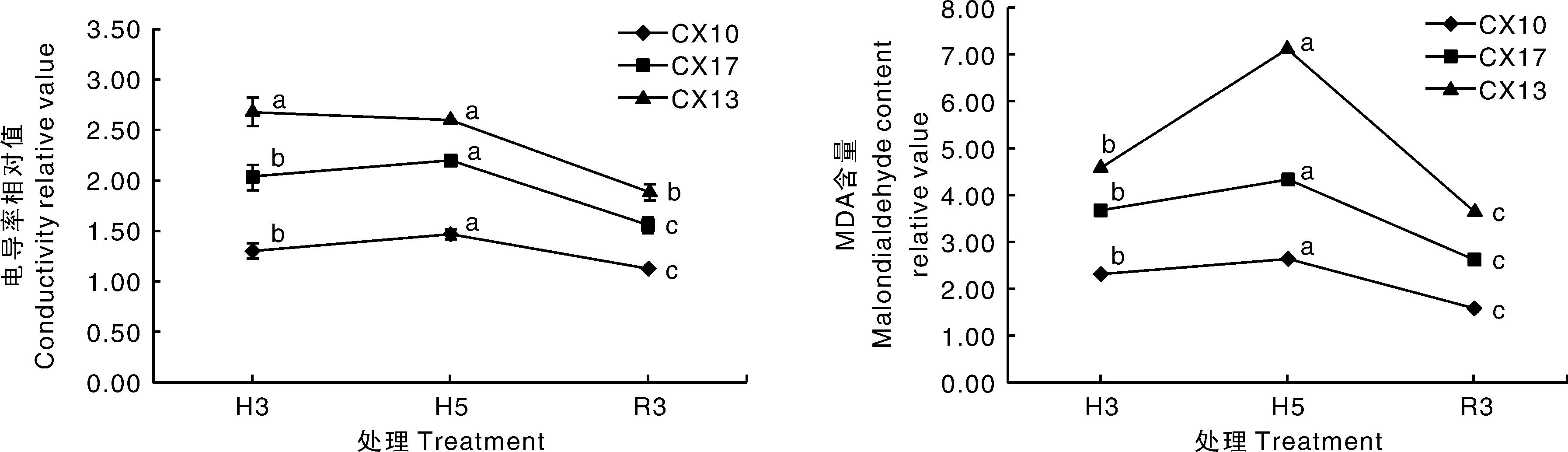
不同处理间没有相同小写字母表示差异显著(P<0.05),下同。Different lower-case letters in the different treatments showed significant differences (P<0.05). The same as below.图3 高温胁迫及恢复对菜心电导率和丙二醛含量相对值的影响Fig.3 Effects of heat stress and recovery on the relative values of conductivity and MDA content in Brassica parachinensis L.

表4 高温胁迫及恢复对菜心SOD、POD、CAT活性相对值的影响Table 4 Effects of heat stress and recovery on the relative values of SOD, POD and CAT activities in Brassica parachinensis L.
同一指标同行数据后没有相同小写字母表示差异显著(P<0.05)。
Data of the same index in the same line followed by different lower-case letters showed significant differences (P<0.05).

图4 高温胁迫及恢复对菜心抗氧化酶基因表达的影响Fig.4 Effects of heat stress and recovery on the expression of the antioxidant enzyme genes in Brassica parachinensis L.
2.6 高温胁迫及恢复对菜心抗氧化酶基因表达的影响
高温胁迫时,抗氧化酶基因Mn-SOD、POD和CAT的表达均高于对照,随着胁迫时间的延长,三者表达量开始出现下降,其中3个抗氧化酶基因均在高温胁迫3 d时达到最大。高温处理3 d和5 d时,耐热型品种CX10的Mn-SOD、POD和CAT基因表达量均显著高于不耐热型品种CX13和中等耐热型品种CX17。与高温胁迫时相比,恢复3 d时,三个品种的抗氧化酶基因表达量均显著下降,但仍高于对照。
3 结论与讨论
植物在生长过程中易受到非生物因素胁迫的影响,持续高温可通过影响植物体内的生理过程而抑制其正常的生长发育[13],植株叶片的形态变化是这种抑制作用最直观的体现。本研究根据菜心热害症状分级,计算热害指数,采用聚类分析法对19个菜心品种进行了耐热性鉴定,获得耐热型品种5个,中等耐热型品种11个,不耐热型品种3个。菜心为喜冷凉蔬菜,当遇到高温时,其生长发育就会受到影响。
正常情况下,细胞内活性氧的产生和清除体系处于动态平衡,此时活性氧水平很低,不会对细胞造成伤害[14]。但当植物受到外界胁迫时,细胞中的活性氧会大量累积,这个平衡就会被打破。SOD、POD和CAT是植物体内酶清除系统中的重要保护酶[15-16],它们的存在可有效清除活性氧自由基,以减轻植物体的受伤害程度。因此,抗氧化酶活性的强弱也常常被认为是衡量植物耐热性的重要指标[17-18]。本研究表明,随着高温胁迫时间的延长,菜心叶片中电导率、MDA含量均呈上升的趋势,其中耐热性强的菜心较对照增幅较小,耐热性差的菜心增幅较大。这主要是由于高温胁迫过程中,耐热性强的菜心叶片保护酶活性迅速升高,并迅速积累活性氧清除剂,如SOD、CAT和POD活性显著高于不耐热品种,前人在甜瓜[19]、甜椒[20]和芥蓝[21]上也有相似的报道。这说明菜心植株可通过协调体内保护酶活性的动态平衡,来增强抗热性,从而减轻高温带来的伤害,这是其对高温胁迫的适应性反应和耐热生存的重要调节机制。胁迫恢复3 d时,植株叶片中电导率、MDA含量、抗氧化酶活性、抗氧化酶基因表达量较高温胁迫时来说均有所降低,但仍高于对照,说明了高温胁迫导致菜心植物细胞受到破坏,并不能完全恢复,菜心幼苗可通过自身的调节机制提高抗氧化酶活性,以适应逆境,这有利于植株在经受更高温度或更长时间的高温胁迫时维持较高的耐热性。
Mn-SOD、POD和CAT基因表达量研究表明,高温胁迫时,各处理菜心中Mn-SOD、POD和CAT基因的表达量均升高,随着胁迫时间的延长,各基因的表达略有降低但始终高于对照。胁迫恢复3 d时,与高温胁迫时相比,三者的表达量均有所降低但仍高于对照。从下降幅度来看,各处理下降幅度与参试品种耐热性强弱关系密切,即耐热性强的品种下降幅度较小,耐热性弱的品种下降幅度较大。这与抗氧化酶SOD、POD和CAT活性变化趋势一致。已有研究证明,植物的抗氧化酶基因表达量上升,可提高其对盐[22]、低温[22]和干旱[23]等逆境的耐受能力。说明耐热型品种抗氧化酶活性的提高主要是通过增加Mn-SOD、POD和CAT基因表达量来实现的。
综上所述,高温胁迫造成了菜心电导率和MDA含量的增加。耐热性强的菜心可通过提高Mn-SOD、POD和CAT基因表达量来增强保护酶活性来降低电解质渗漏率和膜脂过氧化水平,而不耐热的菜心保护酶活性升高较慢,恢复水平和能力较差。本研究结果为菜心耐高温机制研究和菜心耐高温育种提供了一定的理论基础。
