稻米锌含量的遗传分析与基因克隆研究进展
2020-03-05吴婷李霞黄凤林胡标林
吴婷李霞黄凤林胡标林*
(1江西省农业科学院水稻研究所/水稻国家工程实验室,南昌330200;2湖南省水稻研究所/农业农村部长江中下游籼稻遗传育种重点实验室,长沙 410125;第一作者:kuaileting1989@126.com;*通讯作者:hubiaolin992@126.com)
锌是人体和水稻生长发育必需的微量元素,具有重要的生理功能,是人体内多种酶活性催化的辅助因子和蛋白质的结构元件,直接参与核酸、蛋白质等生物大分子合成、骨骼发育、能量代谢及氧化还原过程。缺锌(Zn)是人类最易遭受的3种微量营养素(Zn、Fe和VA)缺乏症之一[1],Zn缺乏会导致发育延缓、免疫力下降、糖尿病等慢性病[2]。因此,锌缺乏相关问题日渐引起社会各界的极大关注。
锌在人体内无法自行合成,必须从膳食中摄取。锌在海鲜、动物内脏等动物类食物中最高,全谷类食物中较低,特别是精制谷类食物中含量更低[3]。稻米是全球超过50%人口的主食,是东南亚和南亚人们营养和能量的主要来源[4-5]。据国际水稻研究所(IRRI)报道,精米中的锌含量平均为12 mg/kg,而根据中国营养学会推荐的大米日摄入量,其中锌含量仅为 3.6~6.0 mg,这远低于WHO和FAO每日建议膳食摄入量12~15 mg的标准[6],从而导致了以禾谷类作物为主食的国家和地区锌营养不良现象普遍发生。
据报道,全世界超过20亿人口存在不同程度的“隐性饥饿”,即缺乏锌、铁等矿质元素和维生素,每年近40万的儿童因患锌缺乏症而死亡[7],61%的亚洲人群存在锌摄入量不足的风险,且该趋势仍在不断加剧,锌已成为影响人类健康的全球性重要因素[2]。解决锌缺乏问题传统的策略是使用保健品、强化食品和微量元素补充剂,但难以满足庞大的锌缺乏群体,尤其是低收入人口的现实需求[8]。由于稻米消费量巨大,其锌含量略微提高将有助于缓解人体锌缺乏问题[9-10]。其次,稻米Zn含量与其生物有效性极显著相关[11]。因此,提高稻米锌含量及增加其生物有效性利用,是解决缺锌性营养危机最为有效的办法,特别是对我国的低收入群体意义重大。
因此,开展稻米锌含量遗传、QTL定位及其基因功能等方面的研究,具有重要理论价值和现实意义。本文综述了稻米锌含量的遗传变异、分布模式、遗传模式、QTL定位和基因克隆等方面研究进展,以为稻米锌含量遗传育种提供参考和依据。
1 稻米锌含量不同基因型的遗传差异性研究
由于育种选择和遗传进化等因素,稻米Zn含量在不同基因型种质间存在显著的遗传差异性,籼稻稻米Zn含量普遍高于粳稻[4]。不同报道中的糙米Zn含量不同,有报道认为是在 13.5~58.4 mg/kg 之间[10],也有人认为是在 15.7~65.0 mg/kg 之间[12],表明通过遗传改良提高稻米Zn含量是可行的。PINSON等[12]对全球1 763份水稻种质在淹水和非淹水栽培模式下糙米中Zn等16种矿质含量的研究表明,非淹水下Zn等元素含量的遗传变异高于淹水下的遗传变异,从而说明糙米锌含量遗传变异不仅由基因型差异决定,还受环境因素的影响。为了了解不同水稻类型的糙米矿质含量的遗传变异性,曾亚文等[13]测定了653份云南稻种资源的糙米锌含量,表明糙米Zn含量的遗传多样性粳稻要高于籼稻。
2 锌在稻米不同部位的分布模式
稻米可分为糙米和精米,精米是加工过程中糙米除去8%~10%麸皮(主要为种皮和胚)的剩余部分[14]。由于饮食习惯和口感,人们日常食用的稻米主要为精米,尽管其营养价值低于糙米。因此,了解锌在稻米不同部位的含量及分布模式非常必要。
研究发现,精米Zn含量为糙米的70%~82%[14-18],可能与其在稻米中转运和分布模式密切相关。在水稻颖果发育过程中,胚乳的表层向内至数层细胞会形成糊粉层组织,主要涉及转移母体组织的脂质、蛋白质和矿质元素等灌浆营养物质至胚和胚乳中积累[19]。Zn从稻米的糊粉层到胚乳均有分布[20]。而HANSEN等[21]研究表明,在加工过程中精米Zn元素的损失量相差也高达5倍。
3 稻米锌含量的经典遗传研究
锌含量等稻米营养品质遗传同时受胚、胚乳、母体植株基因以及细胞质的影响。ZHANG等[22]利用种子胚乳数量性状的遗传模型分析糙米锌含量的遗传效应,表明受种子效应、母体效应和细胞质效应控制,种子加性效应起主要作用,狭义遗传力较高;赖来展等[23]分析了稻谷中锌含量的配合力效应和遗传模型,结果表明,锌含量受加性和非加性效应控制,以加性效应为主,表现为加性-显性模型。张名位等[24]利用7份材料进行完全双列杂交,研究配合力和稳定性,表明糙米锌含量受加性效应和非加性效应控制,以加性效应为主,且还受环境因子影响,表现为基因型与环境互作。可见,稻米锌含量主要受加性和非加性效应控制,以加性效应为主,基因间存在上位性效应,同时又受到微效多基因的影响,属于多基因控制的数量性状。
4 稻米锌含量的QTL定位
在水稻中总共检测到99个Zn含量QTL,包括糙米Zn含量QTL 85个和精米Zn含量14个(见表1)。这些QTL广泛分布于水稻的12条染色体上,其中第8染色体最多,表型贡献率介于2.8%~35.4%之间,其中主效QTL达55个。
在第1染色体长臂和短臂上分别检测到6个和1个Zn含量QTL中,其中HUANG等[25]检测到糙米Zn含量微效QTL qZn1-1位于短臂上,贡献率为6.9%;而长臂上的6个稻米Zn含量QTL成簇分布相似的染色体区域[25-32],其中YU等[32]检测到qZn1控制精米Zn含量,DU 等[26,29]检测的 QTL 为主效 QTL,贡献率分别为14.0%和15.0%。在第2染色体长臂上的9个糙米Zn含量QTL中,黄莹莹等[33]定位到的Zn含量QTL qZn-2,表型贡献率最大,为35.4%。在第3染色体长臂和短臂上检测到的7个和3个Zn含量QTL,其中短臂上3个糙米Zn含量QTL位于同一染色体区域[30,34-35],而ANURADHA等[34]鉴定到的qZn3.1表型贡献最大,为31.0%;在长臂上的 7 个 Zn 含量 QTL 中,YU 等[32,36]检测的主效QTL控制精米Zn含量,其余5个QTL控制糙米Zn含量[5,26,30]第4染色体长臂上检测到的6个糙米 Zn 含量 QTL 成簇分布相似的染色体区域[25,31,35,37-39],其中,HUANG 等[25,35]鉴定到的 QTL 为微效 QTL,其余 4个QTL均为主效QTL。在第5染色体长臂和短臂上分别检测到3个和5个Zn含量QTL。短臂上的3个稻米Zn含量QTL位于相同染色体区域,其中qZN-5为控制精米Zn含量的主效QTL[40],另2个QTL控制糙米Zn 含量[31,41];长臂上 5 个 QTL 中,黄莹莹等[33]鉴定到的糙米Zn含量qZn-5为主效QTL;其余4个QTL为微效 QTL,且成簇分布相似染色体区间[25,32,39],其中 qZn5控制精米Zn含量[32]。在第6染色体的长臂和短臂上分别检测到6个和8个Zn含量QTL,短臂上的8个QTL成簇分布于相似染色体区域[27,28,32,37-39,42-43],其中 qZn6控制精米Zn含量,其余7个QTL控制糙米Zn含量。在长臂上6个QTL位于相似的染色体区域,其中qZn6控制精米Zn含量[36],其余QTL控制糙米Zn含量[30-31,35,37]。在第7染色体的长臂和短臂上分别检测到8个和1个Zn含量QTL,沈希宏等[5]和NORTON等[27]分别检测到1个主效Zn糙米含量QTL,位于长臂相同染色体区域;而 3 个精米含量 QTL[35,36,40]和 3 个糙米 Zn 含量[34,39]成簇分布于长臂上的另一染色体区域,ANURADHA等[34]在染色体短臂上检测到的糙米Zn含量QTL qZn7.3表型贡献率为29.0%。在第8染色体的共鉴定到8个Zn含量QTL,其中4个糙米Zn含量QTL[26,27,30,41]和 1 个精米 Zn 含量 QTL[36]成簇分布于短臂上的相同区域;而SWAMY等[30-31]分别定位到1个糙米Zn含量QTL位于长臂上的相同染色体区域。在第9染色体长臂上相同区间共检测到6个糙米Zn含量QTL,其中黄莹莹等[33]报道的QTL表型贡献率最高,为24.1%,SWAMY 等[30]报道的 QTL 表型贡献率最小,只有4.0%。在第10染色体长臂和短臂上分别检测到4个和3个Zn含量QTL,其中3个糙米Zn含量QTL位
于短臂上的相同区域[30,39]。NORTON等[27]在长臂上检测到2个主效的糙米Zn含量,其中1个QTL qZn10.2的染色体区域分别与1个糙米Zn含量QTL[44]和1个同时控制糙米和精米Zn含量QTL[35]相同。在第11染色体上共鉴定到5个Zn含量QTL,其中,YU等[32-41]分别检测到的主效和微效QTL位于相同染色体区域,均控制精米Zn含量,其余3个QTL控制糙米Zn含量[28,31,42]。第 12 染色体共鉴定到 9 个糙米 Zn 含量 QTL 和1个精米Zn含量QTL,其中XU等[36]检测的精米Zn含量QTL qZn12与ANURADHA等[34]报道的糙米Zn含量QTL qZn12.1的染色体区域相同。另外8个糙米Zn含量QTL中,SWAMY等[30,35]检测到的Zn含量 QTL位于染色体短臂上的相同区域,且其增效等位基因来自野生稻;而 STANGOULIS 等[29,33,41]共同检测到 Zn 含量QTL位于长臂上的相同区域。上述QTL报道中的24套遗传群体主要为籼籼交、籼粳交和粳粳交组合,其中5 套为野栽交群体[30,35,41,44],表明野生稻稻米 Zn 含量基因发掘及QTL研究相对薄弱。不同研究中稻米Zn含量QTL的染色体区间和遗传效应差异大,不仅受遗传组合、群体大小、分子标记、遗传图谱密度及统计方法等影响,可能还与遗传异质性有关,说明稻米矿质遗传机制非常复杂。其次,这些QTL的准确率、作图效率和表型贡献率均较低,只是染色体上一个较大的置信区间。
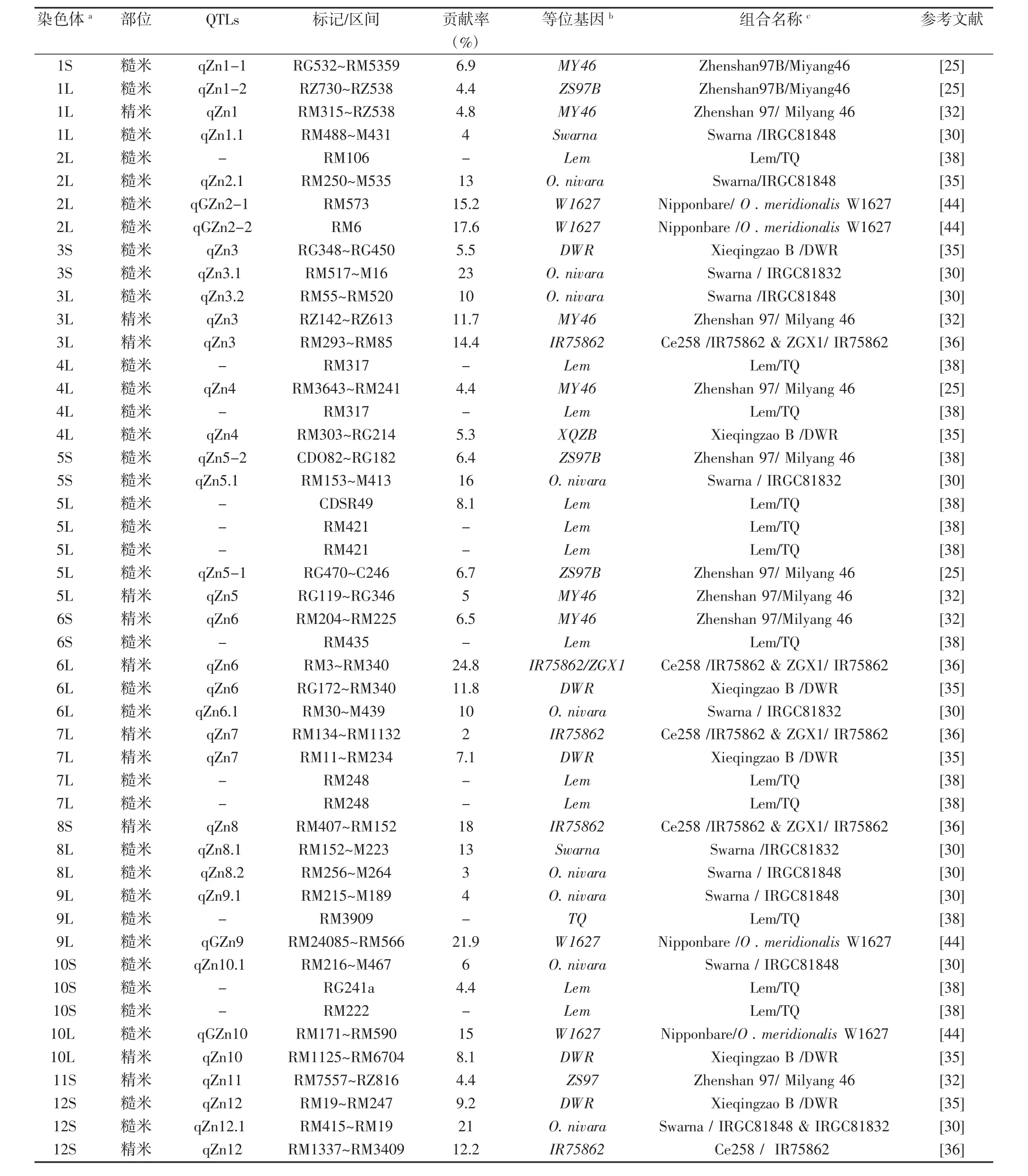
表1 部分文献内容
因此,进一步精细定位尤为必要。YU等[32]将第6染色体短臂上的稻米Zn含量QTL qZn6精细定位至区间 RM19410~Si2944,物理位置约 29.9 kb,包含 3 个注释基因。ISHIKAWA等[44]将第9染色体上稻米Zn含量QTL qGZn9a精细定位至 190 kb区间 RM24211~RM24218,含8个推定基因,其中Os09g0384900是一个强有力的候选基因。ANURADHA等[34]从重组自交群体中检测控制Zn含量QTL,筛选出11个Zn含量高于80 mg/kg的株系,其中株系176(M)中提高Zn含量的QTL最多,其糙米Zn含量为94 mg/kg。上述研究为将来稻米Zn含量分子育种提供了有益的参考。
同时,很多研究发现,稻米Zn与不同矿质元素含量具有相同的QTL。如DU等[26]检测到的控制糙米Zn含量QTL区间RM1111~SSIII2共同影响糙米K、Mg、Fe和Mn含量,这将有助于通过标记辅助选择来协同提高稻米中不同的矿质含量。另一方面,稻米Zn与Cd、As和 Pb等重金属含量具有相同的 QTL,如ZHANG等[59]对16种糙米元素含量进行QTL分析,共检测134个QTL,在8个控制糙米Zn含量QTLs中有3个与控制Cd、As和Pb等重金属含量的QTLs连锁;类似地,XU 等[36]检测了 22个精米 Fe、Zn、Cd和 Pb含量的QTL中,9个控制精米Zn、Cd和Pb含量QTL分别分布于4个基因重叠区域(GO-I–VI):其GO-I包含 qZn3 和 qCd3b,GO-III包含 qZn6 和 qCd6,GO-IV包含qZn7和qPb7,GO-V 包含qZn8、qCd8和qPb8。这些Zn含量与重金属含量QTL研究可为标记辅助选择方法来针对性的选择Zn含量QTL,消除重金属或植酸抗营养因子QTL的不利连锁,精准实现Zn含量分子设计育种具有指导意义。
5 稻米锌含量相关的基因克隆
稻米锌含量的高低与水稻基因型、环境因子(土壤质地、肥力水平、pH值等)密切相关,涉及吸收、转运、累积等信号转导,基因调控等复杂机制,导致基因克隆的复杂与困难。然而随着生物技术和生物信息学发展,相继获得了一批稻米Zn含量相关基因。
目前,已经发现多个与Zn吸收、转运和积累机制相关的基因,如烟酰胺合成酶基因家族(OsNAS1、Os-NAS2、OsNAS3)、 寡 肽 转 运 基 因 家 族(OsPT2、OsYSL15)、ZIP(ZRT/IRT相关蛋白)基因家族(OsZIP1、OsZIP3、OsZIP4、OsZIP5、OsZIP7、OsZIP8)、液泡膜转运蛋 白 (OsVIT1、OsVIT2)、OsFER2、OsIRT1、OsMTP1、OsHMA2和OsFRDL1等[8-9,45-46]。JOHNSON 等[47]研究发现,水稻NAS基因OsNAS1、OsNAS2和OsNAS3的过表达可显著提高糙米Zn含量。其中,OsNAS3基因上调表达不仅可以增强植株对Zn缺乏的耐受力,还能忍受过量Zn元素的胁迫,其转基因植株籽粒中Zn含量提高了2.3倍。LEE等[48]报道,过表达OsNAS2可提高精米Zn 含量 2.4~2.9 倍。PAUL 等[49]发现,过表达水稻铁蛋白基因OsFER2提高锌含量1.4倍。TAKAHASHI等[50]构建了OsSUT1启动子控制的OsHMA2过表达载体转化水稻,结果发现,精米Zn含量得到少量的提高。ZIP家族参与Zn2+转运,OsZIP1和OsZIP3主要负责从土壤中吸收 Zn,OsZIP4、OsZIP5和OsZIP8主要负责 Zn由根部向茎部的运输,而OsZIP4和OsZIP8主要负责Zn转运至稻谷中。ISHIMARU等[51]将由CaMV35S作为启动子的OsZIP4基因过表达载体导入水稻中,发现转基因水稻根部的Zn含量是对照组的10倍,但其茎秆中Zn含量减少到对照组的1/5,籽粒中Zn含量也减少为对照组的1/4。LEE等[52]将OsZIP5基因的过表达载体转入水稻中,发现OsZIP5的表达模式及功能与OsZIP4相似,OsZIP5过表达水稻根部锌大量积累,而其地上部分的Zn含量较对照组低。LEE等[53]对OsZIP8转基因水稻锌元素分布的研究也证实,过表达OsZIP8基因可促进根细胞对Zn的吸收并干扰Zn向地上部分运输,导致植株出现秸秆变矮、籽粒减产等Zn缺乏的症状。
6 富锌水稻的育种研究
微营养元素Zn的缺乏是导致人体“隐形饥饿”最重要的因素之一,稻米Zn含量的提高有助于缓解人体锌缺乏的问题。合理广泛鉴定富Zn的水稻种质资源是高锌水稻育种的基础。
ANURADHA等[34]应用重组自交群体进行糙米Zn含量QTL分析,从中筛选出11个Zn含量高于80 mg/kg的株系,其中株系176(M)中携带Zn含量的QTL最多,而其糙米Zn含量高达94 mg/kg。KUMAR等[54]从Palman579和PAU201杂交F2群体中筛选得到Zn含量高达157.4 mg/kg的优良系。雷国方等[55]利用功亲1号和功亲2号杂交,选育出紫米新品种功米2号,其Zn含量有77.90 mg/kg,是普通稻米的2倍多。吴敬德等[56]等从国际水稻研究所引进5份富Zn水稻种质IR41994-50-2-1-3(S1)、IR62141-114-3-2-2-2(S2)、IR63429-23-1-3-3(S3)、Rexmont(S4) 和 IR68144-213-2-2-3(S5),其精米Zn含量较常规水稻要高,而且其中的S1和S5的Fe含量也高于常规水稻,可以作为富Zn水稻种质资源亲本开展高Zn水稻品种选育。张琳琳等[57]利用Co60-γ射线辐射诱变YN56获得高Zn突变体,经过多代获得富Zn新品系——粒粒滋,经检测在杭州和余姚种植收获的籽粒精米Zn含量分别高达32.8 mg/kg 和 34.1 mg/kg。
7 展望
相对于保健品、补充剂、药剂等,人们更倾向于选择富含微量元素且不改变外观、口感和烹调特性的生物强化作物[58]。锌缺乏已成为影响人体健康的严重的公共卫生问题,因此,解决锌缺乏症是国际Harvest Plus计划中最重要的研究目标。通过生物强化手段来提高水稻对锌的吸收、转运和积累进而提高稻米锌含量,成为降低全球禾谷类食用人群的锌缺乏症的策略。
不同基因型水稻对锌元素的吸收与利用存在显著差异,且水稻籽粒中的锌含量与胚乳中的锌含量无直接关系,糙米中的锌生物有效性又受抗营养因子植酸的影响。有关水稻锌吸收、转运和积累机制相关的IRT1基因、HMA基因、ZIP基因、NAS基因、OsVIT1和Os-VIT2基因的克隆研究,多数与铁、重金属转运蛋白和多种离子运输相关,直接与锌相关的较少,且多数基因以在植株其他部位表达为主,应加强开展稻米尤其是胚乳中锌转运基因的克隆。因此,全面深入探究稻米Zn含量分子机理,为水稻育种及分子设计育种提供重要参考,具有重要的理论价值与现实意义。
