基于高通量测序技术的两种水螅水母现场食物研究
2020-02-28徐盛楠孙婷婷彭赛君张建设董志军
徐盛楠,孙婷婷,彭赛君,张建设,董志军,3
(1.浙江海洋大学国家海洋设施养殖工程技术研究中心,浙江 舟山 316022;2.中国科学院烟台海岸带研究所牟平海岸带环境综合试验站,山东 烟台 264003;3.中国科学院海洋大科学研究中心,山东 青岛 266071)
水母以摄食量高、生长速度快、食物来源广泛等特征,在海洋食物网中占有十分重要的地位[1]。近年来,水母暴发事件频发,已成为近岸海域典型的海洋生态灾害[2-3]。暴发的水母类需要消耗大量食物,比如浮游生物、鱼虾贝类幼体等,而激增的摄食量对被摄食者的种群数量和分布都具有不可忽视的影响。同时,水母又可被多种生物类群摄食,企鹅(Spheniscus)、银鲳(Pampusasgenteus)、绿鳍马面鲀(Navodonseptentrionalis)以及大型海龟棱皮龟(Dermochelyscoriacea)等生物类群都会摄食一定数量的水母作为其能量补充[4-7]。因此,进行水母类的摄食研究对探究水母在海洋食物网中的物质循环及能量流动的地位具有重要意义。
目前,国内外学者对水母类摄食已有较多研究[8-12],采用的方法主要包括直接镜检法、肠道色素法、脂肪酸标记法和同位素标记法。其中,直接镜检法是最常见的生物食性检测方法,但该方法难以从海洋生物胃腔中鉴定半消化食物种类,同时,摄入的食物种类通常缺乏清晰的分类学诊断特征,不适用于个体较小生物的食物种类鉴定[13-14];肠道色素法只能测定含有色素的植物摄食物,无法有效测定不含色素的动物及其碎屑[15];脂肪酸标记法可以揭示食物网中的物质流动,定量分析各类食物来源对摄食者的食物贡献程度,但易受到生物特定脂肪酸在空间和时间上的变化[16]、特定脂肪酸在捕食者体内的变化[17]及脂肪酸的降解[18]等因素的限制;同位素标记法能够以碳氮同位素在生物之间的稳定富集作用来揭示捕食者的食物组成,但该方法在复杂的食物网中难以获得更为准确的食物种类结果[19]。
高通量测序技术(High-throughput DNA Sequencing Technique)是现代分子生物学最重要的分析方法之一,具有信息量大、灵敏度高、成效快、成本低等优势,越来越广泛地应用于生物多样性的研究[20]。目前,高通量测序技术已被应用于鱼类和龙虾幼体等海洋生物的摄食多样性研究中,揭示了丰富的摄食生物多样性[21-24]。本研究利用高通量测序技术来研究管花萨氏水母(Sarsiatubulosa)和八斑芮氏水母(Rathkeaoctopunctata)这两种我国北方近海沿岸水域常见的水母种类的现场食物组成,为快速准确地检测水母类现场食物组成提供新的技术方法。
1 材料与方法
1.1 样品采集和处理
2018年4月,于威海荣成近岸海域(36.934°N,122.419°E)采用浅水Ⅰ型浮游生物网(网口直径50 cm,筛绢孔径0.505 mm)拖网采集管花萨氏水母和八斑芮氏水母,分装于15 cm3的灭菌离心管中,并加入2%的中性鲁哥氏液,避光保存。每个离心管中放置30只管花萨氏水母,共12个样品;10只八斑芮氏水母,共12个样品,总计24个样品。现场固定的水母样品带回实验室后,4 ℃下避光保存。DNA提取前,使用经0.22 μm滤膜过滤的无菌海水清洗样品表面2~3次,并在OLYMPUS SZX10体视显微镜下观察,确保清洗之后的管花萨氏水母和八斑芮氏水母表面无其他生物或碎屑附着,处理好的样品吸干水分后,转移到1.5 cm3的灭菌离心管中,每个离心管放置1只水母样品。
1.2 样品DNA提取
将两种水母样品置于-80 ℃超低温冰箱冷冻30 min,用灭菌过的杵棒充分研磨样品。加入500 mm3DNA裂解液,于恒温水浴锅中55 ℃水浴48 h,样品总DNA提取后,使用GeneJET Gel Extraction Kit纯化试剂盒对DNA样品进行纯化,纯化后的DNA样品冷冻保存于-20 ℃冰箱。
1.3 PCR扩增、文库构建与高通量测序
使用带Barcode特异引物的18S V4区528F-706R为引物对提取的DNA进行PCR扩增。引物序列:528F:5′-GCCTCCCTCGCGCCATCAGGCGGTAATTCCAGCTCCAA-3′;706R:5′-GCCTTGCCAGCCCG ATCAGAATCCRAGAATTTCACCTCT-3′。30 mm3的反应体系包括15 mm3Phusion Master Mix,3 mm3引物,10 mm3DNA 和2 mm3ddH2O。在Bio-rad T100梯度PCR仪上进行PCR扩增反应:98 ℃预变性1 min;98 ℃变性10 s,50 ℃退火30 s,72 ℃延伸30 s,30个循环;最后72 ℃保温5 min。使用GeneJET胶回收试剂盒(Thermo Scientific)纯化PCR产物,根据PCR产物浓度进行等质量混样操作,充分混匀后用2%的琼脂糖凝胶电泳纯化PCR产物,选择主带大小在400~450 bp的序列,割胶回收目标条带。使用Illumina公司Truseq DNA PCR-Free Library Preparation Kit建库试剂盒进行文库的构建,构建好的文库经Qubit定量和文库检测,合格后在Illumina HiSeq PE250平台上对文库进行测序,并产生300 bp的配对末端读数。
1.4 数据分析
对测序所得到的原始数据(Raw Data)进行拼接和过滤,获取有效数据(Clean Data)。利用获得的有效数据进行OTUs(Operational Taxonomic Units)聚类和物种分类分析,将所得物种18S rDNA的序列在GeneBank上找到对应的物种,利用MEGA 6.0软件构建Neighbor-joining(N-J)生物系统进化树进行聚类分析,利用SPSS 19.0软件对水母的各门类和纲类摄食物种进行对比分析,得到两种不同水母的摄食结构。
2 结果与分析
2.1 实验结果
由Illumina HiSeq PE250平台测序后,本研究共获得原始序列2 188 641段,经拼接和过滤后得到有效序列为2 124 548段,共筛选有用信息共34个OTUs用于进一步分析。管花萨氏水母和八斑芮氏水母的具体食物组成及占比具体见表1。
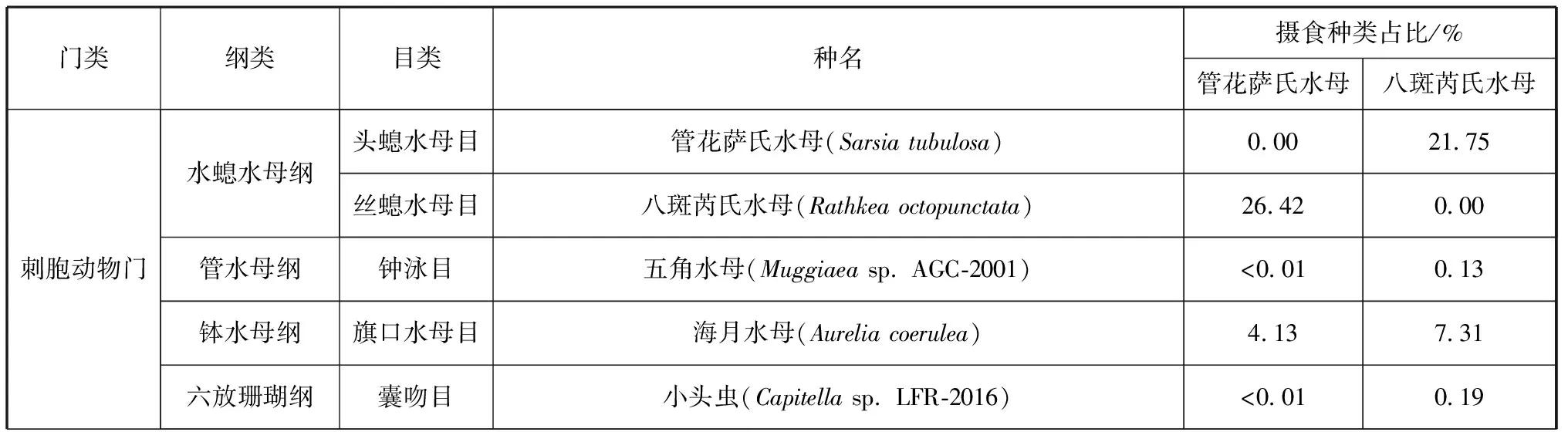
表1 管花萨氏水母和八斑芮氏水母食物组成及占比
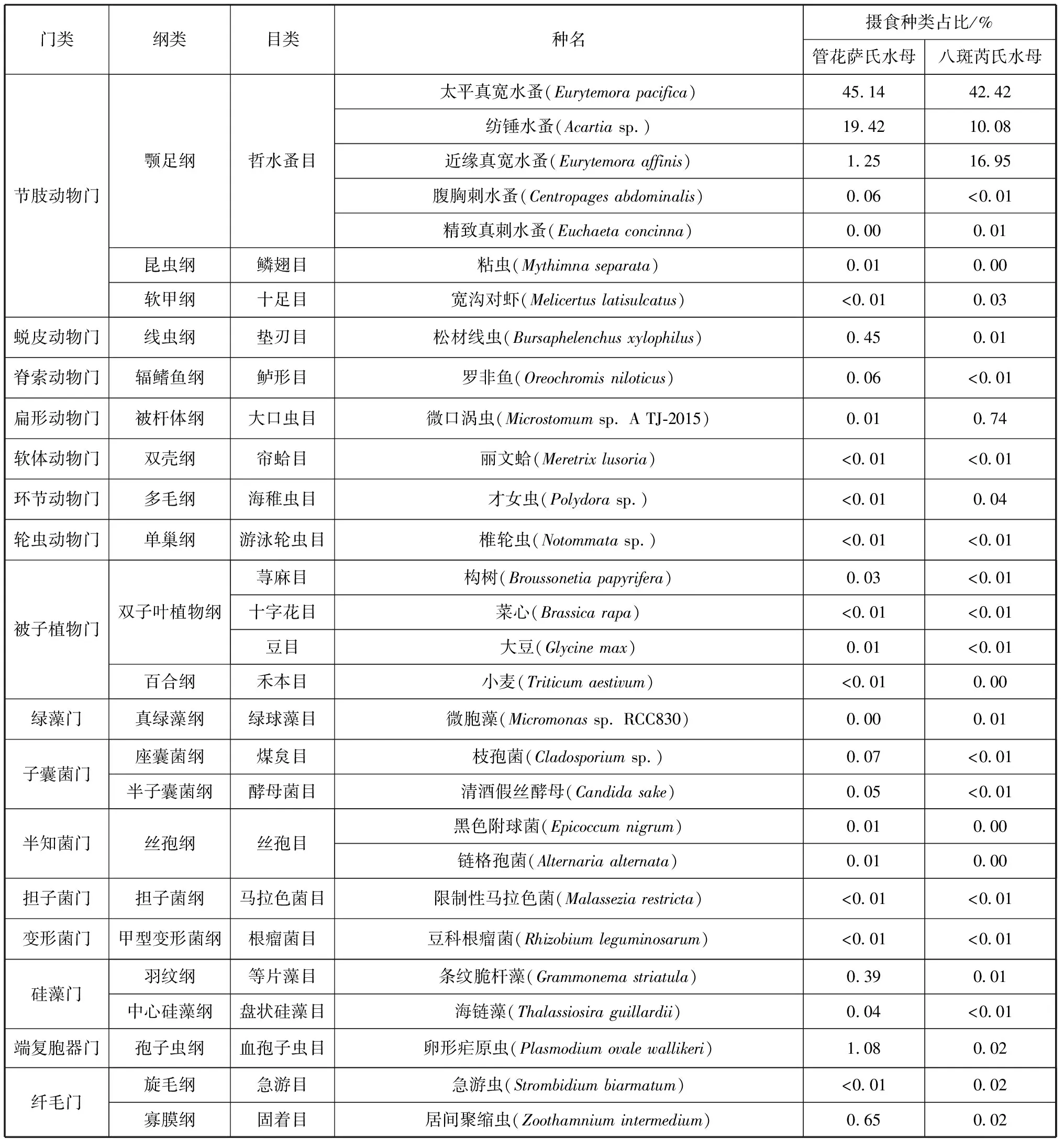
续表1
两种水螅水母共鉴定出34种食物种类,其中管花萨氏水母食物种类为31种,包括刺胞动物门、节肢动物门、端复胞器门、纤毛门等16个门类,颚足纲、钵水母纲、孢子虫纲、寡膜纲等25个纲类。管花萨氏水母现场摄食食物的主要门类组成为节肢动物门和刺胞动物门,分别占其食物来源的66.29%和30.76%(图1);主要的纲类组成为颚足纲、水螅水母纲和钵水母纲,分别占其食物来源的66.28%、26.58%和4.16%(图2)。管花萨氏水母主要的食物种类是太平真宽水蚤(Eurytemorapacifica)和八斑芮氏水母,二者分别占到其食物来源的45.14%和26.42%(表1)。
八斑芮氏水母共鉴定出29种食物种类,包括节肢动物门、刺胞动物门、扁形动物门、纤毛门等16个门类,颚足纲、水螅水母纲、钵水母纲、被杆体纲等23个纲类。八斑芮氏水母现场摄食食物的主要门类组成同样为节肢动物门和刺胞动物门,分别占69.63%和29.44%(图3);主要的纲类组成为颚足纲、水螅水母纲和钵水母纲,分别占69.59%、21.79%和7.31%(图4)。八斑芮氏水母主要的食物种类是太平真宽水蚤和管花萨氏水母,分别占42.42%和21.75%(表1)。
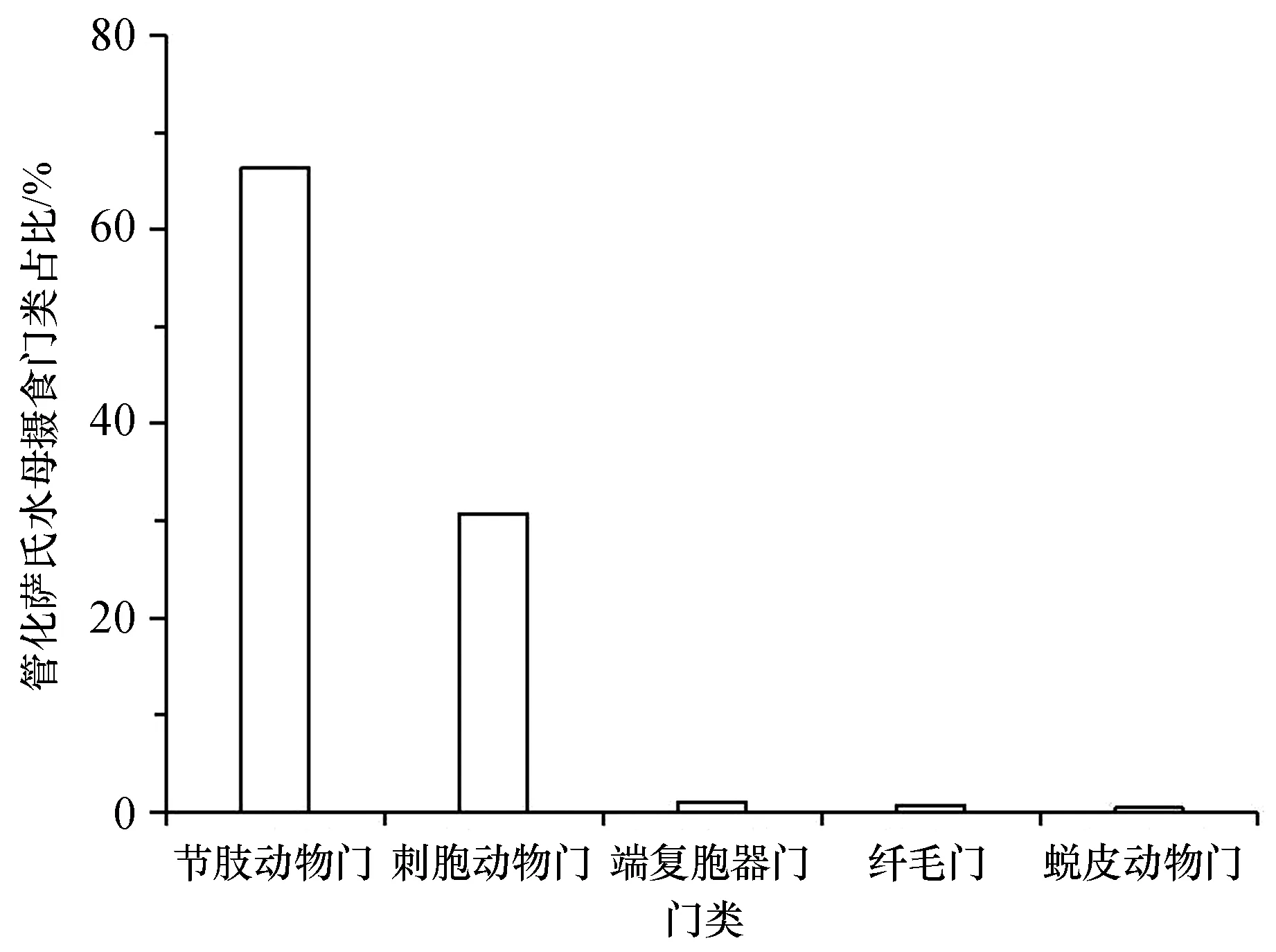
图1 管花萨氏水母现场摄食食物种类的主要门类占比
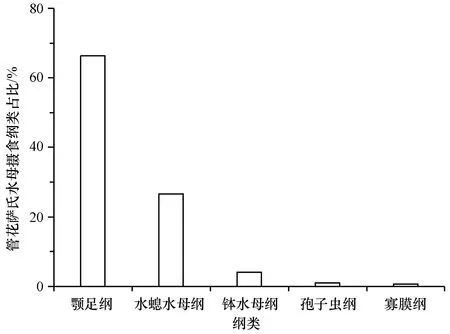
图2 管花萨氏水母现场摄食食物种类的主要纲类占比
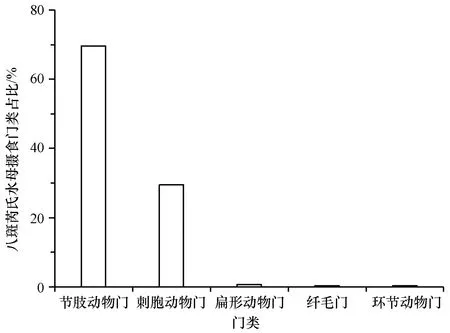
图3 八斑芮氏水母现场摄食食物种类的主要门类占比
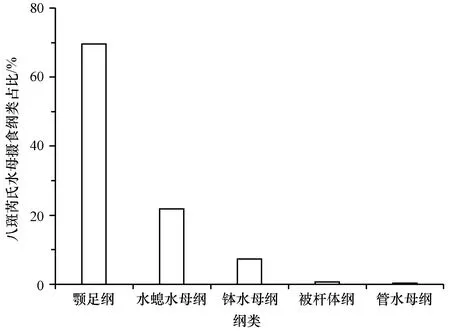
图4 八斑芮氏水母现场摄食食物种类的主要纲类占比
基于18S rDNA,使用邻接法构建的Neighbor-joining(N-J)生物系统进化树(图5),对管花萨氏水母和八斑芮氏水母的现场摄食食物进行的聚类分析发现,两者的共同摄食种类包括海月水母(Aureliacoerulea)、太平真宽水蚤、纺锤水蚤(Acartiasp.)、近缘真宽水蚤(Eurytemoraaffinis)等26种食物,其中桡足类为4种,且该类群在管花萨氏水母和八斑芮氏水母的摄食食物组成中占有很高的比例,分别为66.28%和69.59%。管花萨氏水母独有的摄食食物种类包括八斑芮氏水母、粘虫(Mythimnaseparata)、小麦(Triticumaestivum)、黑色附球菌(Epicoccumnigrum)和链格孢菌(Alternariaalternata)5种,而八斑芮氏水母独有的摄食食物种类则包括管花萨氏水母、精致针刺水蚤(Euchaetaconcinna)和微胞藻(Micromonassp.RCC830)3种。
2.2 讨论
研究结果表明,管花萨氏水母和八斑芮氏水母现场摄食食物种类具有较高的多样性。管花萨氏水母摄食食物种类31种,涉及节肢动物门、刺胞动物门、端复胞器门等16个门类,颚足纲、水螅水母纲、钵水母纲等25个纲类;八斑芮氏水母摄食物种类29种,涉及节肢动物门、刺胞动物门、扁形动物门等16个门类,颚足纲、水螅水母纲、钵水母纲等23个纲类,摄食种类多样,门类广泛。水母的广食性,在本研究中再一次得到了证实,在之前的对水母摄食的研究中,已有研究学者提出此观点。Purcell等(2003)在对海月水母、狮鬃水母(Cyaneacapillata)和多管水母(Aequoreaaequorea)3种大型水母的摄食研究中发现,桡足类、桡足类幼体和枝角类都是它们的主要摄食对象[25]。陈丽华等(2003)发现港球侧腕水母(Pleurobrachiaglobosa)可摄食长尾类幼体、短尾类幼体、蔓足类无节幼体、桡足类和端足类等多种生物种类[9]。Riascos等(2016)发现在大型钵水母Chrysaoraplocamia的摄食物种中鱼卵、鱼幼体和甲壳类浮游动物均占较大比例,约52.4%~99.3%[26]。Underwood等(2007)在伊鲁坎吉水母(Carukiabarnesi)肠道中发现了鱼卵、鱼类的幼体和节肢动物门的动物[11]。由此可见,水母的摄食食物种类多样,且来源广泛。
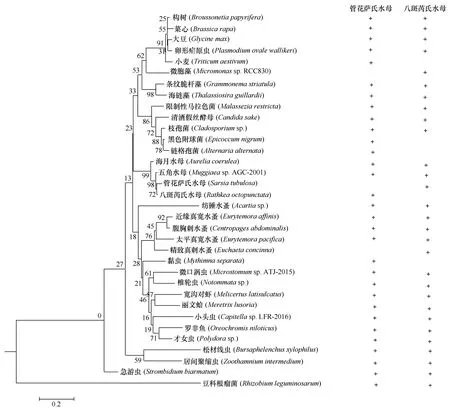
图5 基于18S rDNA构建Neighbor-joining(N-J)生物系统进化树
管花萨氏水母和八斑芮氏水母的的主要食物来源均为节肢动物门、颚足纲,主要食物种类均为桡足类的太平真宽水蚤。Pagès等(1996)对薮枝水母(Obeliaspp.)和八斑芮氏水母的胃含物的检测,以拟长腹剑水蚤(Oithonasimilis)和诺氏三角溞(Evadnenordmanni)两种桡足类生物为主要的摄食物种[27],与本研究得出的主要摄食的桡足类种类不同,推测可能是由于水母所处海域环境不同,桡足类优势种存在差异造成的。在Arai(1997)对霞水母(Cyaneasp.)和口冠水母(Stomolophusmeleagris)的胃含物的分析研究中,同样发现桡足类是其摄食的主要食物类群[28]。因此,桡足类是水母类的重要食物来源。
本研究发现,管花萨氏水母和八斑芮氏水母的摄食食物除浮游动物外,还包括少量真菌、浮游植物。在管花萨氏水母的摄食物种中发现了枝孢菌(Cladosporiumsp.)、清酒假丝酵母(Candidasake)、黑色附球菌、链格孢菌和限制性马拉色菌(Malasseziarestricta)5种真菌以及条纹脆杆藻(Grammonemastriatula)和海链藻(Thalassiosiraguillardii)2种藻类;在八斑芮氏水母的摄食物种中发现了枝孢菌、清酒假丝酵母和限制性马拉色菌3种真菌以及微胞藻、条纹脆杆藻和海链藻3种藻类。因此,管花萨氏水母和八斑芮氏水母均属于杂食性。本研究发现水母摄食组成中含有真菌和浮游植物并非偶然,曾有学者提到前寒武纪时期,在较大型的浮游生物出现之前刺胞动物会摄食真菌和藻类[29]。马喜平等(1998)认为水母会选择性地摄食一些浮游植物和有机碎屑来补充其能量需求[30]。然而真菌和浮游植物与水母之间的营养关系还存在其他可能。真菌在沿海水域极为丰富,通常附着于浮游植物、浮游颗粒和有机聚集体上或以自由形式存在[31]。众多研究发现桡足类的现场摄食食物中含有枝孢菌、马拉色菌(Malassezia)和短梗霉菌(Aureobasidium),但它们与海洋桡足类之间的营养关系并没有得到肯定[32-33]。Riemann等(2010)在欧洲鳗(Anguillaanguilla)肠道含物的DNA检测中发现真菌序列,但考虑到与其他海洋生物间的相互作用,并无法确定二者之间的捕食关系[34]。同理,水母和真菌之间的摄食和营养关系同样也需要进一步研究。此外,在本研究中管花萨氏水母和八斑芮氏水母对浮游植物的摄食比例较小,所摄食的浮游植物以硅藻类占比较大,且硅藻类浮游植物是桡足类摄食的主要食物类群,而桡足类又是管花萨氏水母和八斑芮氏水母主要的摄食物种,由此我们推测成体管花萨氏水母和八斑芮氏水母可能并非直接摄食浮游植物,而是在摄食桡足类时将其体内未消化的浮游植物碎屑一同摄入体内。
研究结果中管花萨氏水母和八斑芮氏水母的摄食食物种类中还含有少量的陆地植物碎屑,包括构树(Broussonetiapapyrifera)、小麦、菜心(Brassicarapa)和大豆(Glycinemax)4种。这4种陆地植物为当地的常见物种,且在采样时间内较为丰富,因此,可能成为水母摄食的一部分。但无法确定管花萨氏水母和八斑芮氏水母是直接摄食陆生植物(碎屑)以维持其食物摄入量作为能量补充,还是由于摄食的桡足类体内含有未被消化的陆生植物碎屑而间接进入水母体内。有研究者认为,当存在异养微生物时,陆地植物的花粉可以刺激浮游动物的生长[35],除花粉碎屑外,其他类型的陆地植物碎屑也会成为浮游动物摄食的一部分[36-37]。胡思敏等(2015)的研究发现陆地植物在桡足类摄食食物中占据一定比例[33]。由于目前尚未见对水母摄食陆地植物和相关物种的研究报道,因此,我们需进行进一步的现场或室内摄食实验分析来了解沿海生态系统中不同生物之间的摄食及营养关系,以确定陆地植物的营养价值和对水母摄食的贡献作用。
管花萨氏水母和八斑芮氏水母互为捕食对象,且在双方食物比重中的占比较高。管花萨氏水母摄食八斑芮氏水母的比例占总摄食食物比例的26.42%,八斑芮氏水母摄食管花萨氏水母的比例也达到了21.75%。除此之外,二者还均摄食海月水母等其他水母种类。水母相互捕食的现象并非个例,Hansson等(1997)发现狮鬃水母会捕食海月水母以及其他水母[38],马喜平等也曾在其研究中指出水母类之间的捕食现象较为普遍[30]。
3 结论
本研究利用高通量测序的方法对管花萨氏水母和八斑芮氏水母的现场食物组成进行了研究,对两种水母的摄食结构进行了系统的讨论。其研究结果表明:
(1)管花萨氏水母和八斑芮氏水母的的摄食食物种类具有多样性,管花萨氏水母摄食种类为31种,涉及16个门类、25个纲类;八斑芮氏水母摄食种类为29种,涉及16个门类、23个纲类。
(2)两种水母从摄食物种上看属于杂食性,其食物组成包括浮游动物、真菌、浮游植物、陆地植物碎屑等。
(3)管花萨氏水母和八斑芮氏水母二者互为捕食对象。
(4)二者的摄食结构具有相似性,共包含26种相同的摄食物种,且均以节肢动物门和颚足纲的食物类群为主,以太平真宽水蚤为优势种。
