玉米大斑病菌和小斑病菌交配型多重PCR检测方法的建立与应用
2020-02-25代玉立甘林滕振勇杨静民祁月月石妞妞陈福如杨秀娟
代玉立,甘林,滕振勇,杨静民,祁月月,石妞妞,陈福如,杨秀娟
玉米大斑病菌和小斑病菌交配型多重PCR检测方法的建立与应用
代玉立1,甘林1,滕振勇2,杨静民3,祁月月4,石妞妞1,陈福如1,杨秀娟1
(1福建省农业科学院植物保护研究所/福建省作物有害生物监测与治理重点实验室,福州 350013;2福建省种子管理总站,福州 350001;3建瓯市农业农村局,福建建瓯 353100;4浙江天丰生物科学有限公司,浙江金华 321000)
【目的】由大斑突脐蠕孢()和玉蜀黍平脐蠕孢()引起的玉米大斑病和小斑病是玉米生产上重要的叶部真菌病害,本研究旨在建立玉米大斑病菌和小斑病菌交配型多重PCR检测方法,为玉米大斑病菌和小斑病菌的交配型田间分布和有性生殖研究提供技术方法。【方法】根据GenBank中已登录的玉米大斑病菌(登录号MAT1-1:GU997138和MAT1-2:GU997137)和小斑病菌(登录号MAT1-1:X68399和MAT1-2:X68398)交配型基因序列,利用Primer Premier 5.0软件设计2种病原菌交配型多重PCR检测特异性引物,采用单因素法对引物的退火温度以及扩增程序中延伸时间和循环数等重要参数进行优化,建立玉米大斑病菌和小斑病菌交配型多重PCR检测方法,并对2种病原菌的交配型多重PCR检测灵敏度和特异性进行检验。同时,对田间采集的129株玉米大斑病菌和194株玉米小斑病菌单孢菌株的交配型进行多重PCR检测,以明确建立的交配型多重PCR检测方法的适用性。【结果】建立的多重PCR方法和设计的交配型特异引物StMAT01-2F/R、StMAT02-3F/R、ChMAT01-3F/R和ChMAT02-2F/R可分别扩增出MAT1-1、MAT1-2型菌株大小为816、132 bp(大病斑菌)与490、136 bp(小病斑菌)的特异性目的条带。25 μL多重PCR扩增体系:2×Multiplex PCR Mix 12.5 μL,引物各10 pmol,DNA模板100 ng,退火温度为57.2℃(大病斑菌)和55.0℃(小斑病菌),35个循环。该多重PCR对玉米大斑病菌MAT1-1、MAT1-2型单孢菌株的检测灵敏度分别为0.1、0.01 ng基因组DNA,而对玉米小斑病菌MAT1-1、MAT1-2交配型的检测灵敏度均为0.1 ng基因组DNA。该交配型多重PCR检测方法对玉米大斑病菌和小斑病菌特异性很强,能够很好地区分玉米大斑病菌和小斑病菌相应的近缘种和14株其他真菌菌株。不同地理来源的玉米大斑病菌和小斑病菌交配型检测结果表明,该多重PCR能够准确地检出129株玉米大斑病菌和194株玉米小斑病菌的交配型,且检测结果与随机抽取的菌株杂交验证结果完全吻合。【结论】构建的玉米大斑病菌和小斑病菌交配型多重PCR检测方法灵敏度高、特异性好、操作简便,能够准确、快速地检测出玉米大斑病菌和小斑病菌的交配型,为玉米大斑病菌和小斑病菌的交配型田间分布和监测及有性生殖研究提供了可靠的技术方法。
玉米大斑病菌;玉米小斑病菌;交配型;多重PCR;灵敏度;特异性
0 引言
【研究意义】由玉米大斑病菌()和小斑病菌()分别侵染玉米叶片引起的玉米大斑病(northern corn leaf blight,NCLB)和小斑病(southern corn leaf blight,SCLB)是世界各玉米产区普遍发生的重要叶部真菌病害[1-3]。玉米大斑病和小斑病在中国东北地区、黄淮海地区以及南方山地等玉米主产区发生严重,一般减产10%—30%,大暴发年份减产50%—100%,严重损害了玉米的产量和品质[4-5]。玉米大斑病和小斑病均为典型的流行性病害,随气流传播的分生孢子可在田间循环侵染,造成病害在短时间内流行暴发[6-8]。此外,这两种病原菌均可在玉米生长后期进行有性生殖,造成病原菌种群内遗传变异频繁、生理分化明显以及高寄主适合度菌株不断出现[9-11],从而致使玉米新品种的抗性迅速丧失,这为我国玉米大斑病和小斑病的长期可持续控制带来了困扰。明确植物病原微生物的交配型是进行病害有性生殖研究的基本前提,因此,建立玉米大斑病菌和小斑病菌交配型快速、准确和操作简便的检测方法具有重要的实践意义。【前人研究进展】研究玉米大斑病菌和小斑病菌的交配型、有性生殖及其有性后代的寄主适合度,揭示病原菌的遗传变异机制[5,9,12],可为制定病害的综合防控措施提供理论参考。然而,植物病原菌不同交配型菌株间无明显的形态学差异,研究者无法通过肉眼或显微观察来准确判断病原菌的交配型。传统的交配型检测方法是将待测菌株与交配型已知菌株进行室内杂交,通过有性子实体的形成与否,来判断待测菌株的交配型,该方法检测周期长,易受外界因素的影响,而且往往由于菌株间交配亲和能力弱而导致检测失败[13-15]。随着分子生物学技术的发展,通过设计特异引物用PCR方法检测子囊菌的交配型已成为国内外研究的热点[16-17]。TAKAN等[18]根据稻瘟病菌()MAT1-1和MAT1-2基因序列设计交配型特异引物,采用常规PCR方法检测稻瘟病菌交配型及其在东非的地理分布;DE MICCOLIS ANGELINI等[19]设计了葡萄灰霉病菌()交配型RT-PCR检测特异引物,用于检测葡萄灰霉病菌菌丝、菌核以及子囊盘时期的交配型;BREWER等[17]在研究葡萄白粉病菌()交配型基因座结构的基础上,建立了葡萄白粉病菌交配型的多重PCR检测方法。此外,交配型的PCR快速检测方法还被成功应用于芸薹叶斑病菌()[20]、大麦云纹病菌()[21]、小扁豆壳二孢疫病菌()[15]、小麦壳针孢叶枯病菌()[22]和除虫菊褐斑病菌()[23]等重要植物病原菌的交配型检测中。玉米大斑病菌和小斑病菌均属典型的异宗配合的子囊菌[24]。目前,玉米大斑病菌和小斑病菌的交配型基因序列已被克隆,两种病原菌均具有一对在核苷酸序列上高度异源的交配型基因座,即MAT1-1和MAT1-2[25-26]。MAT1-1和MAT1-2基因在结构上分别至少含一个-1结构域和高泳动蛋白家族(high mobility group,HMG)结构域[25-26]。通过检测-1和HMG结构域即可准确鉴定异宗配合子囊菌的交配型[16-17]。【本研究切入点】在前人已完成玉米大斑病菌和小斑病菌交配型基因克隆的基础上,根据各自交配型基因序列的保守区设计交配型检测特异引物,构建玉米大斑病菌和小斑病菌交配型多重PCR检测体系,提高交配型检测的效率。【拟解决的关键问题】根据玉米大斑病菌和小斑病菌的交配型基因序列,设计其交配型检测特异引物,单因素法优化多重PCR反应条件,建立快速、准确以及可分别同时检测玉米大斑病菌和小斑病菌交配型的多重PCR检测方法,为玉米大斑病菌和小斑病菌的交配型田间分布和监测以及有性生殖研究提供可靠的技术手段。
1 材料与方法
试验于2018年6月至2019年7月在福建省农业科学院植物保护研究所完成。
1.1 供试菌株
供试玉米大斑病菌近缘种參子凸脐蠕孢(;CGMCC编号:3.13861)、小柄凸脐蠕孢(;3.13862)和芦苇凸脐蠕孢(;3.13863)菌株购自中国科学院微生物研究所中国普通微生物菌种保藏管理中心(CGMCC);玉米圆斑病菌()和小麦根腐病菌()菌株分别由四川农业大学农学院龚国淑教授和安徽省农业科学院植物保护与农产品质量安全研究所汪涛助理研究员惠赠;供试的其他真菌菌株均由福建省农业科学院植物保护研究所植物病害研究室分离保存。玉米大斑病菌和小斑病菌标准菌株的编号和交配型以及其他真菌菌株见表1。
1.2 供试菌株基因组DNA的提取
将稻瘟病菌和其他供试菌株分别在淀粉琼脂培养基和马铃薯葡萄糖琼脂培养基(PDA)平板上28℃黑暗培养5—7 d,用无菌手术刀轻轻刮取培养基表面菌丝置于2 mL无菌离心管中,供试菌株基因组DNA的提取采用改良的CTAB法[27]。DNA样品用30 µL TE缓冲液(10 mmol·L-1Tris-HCl,1 mmol·L-1EDTA,pH 8.0)溶解,核酸定量仪测定各菌株DNA的浓度并用TE缓冲液调节至100 ng·μL-1,DNA样品于-80℃冰箱中保存。
1.3 多重PCR特异性引物的设计
从GenBank网站(https://www.ncbi.nlm.nih.gov/)下载玉米大斑病菌交配型MAT1-1和MAT1-2(登录号:GU997138和GU997137)以及玉米小斑病菌交配型MAT1-1和MAT1-2(登录号X68399和X68398)的基因序列[25-26]。利用DNAMAN 5.0软件进行多重序列比对,寻找各交配型基因的保守序列。通过Primer Premier 5.0软件设计交配型多重PCR检测的特异引物,利用Primer-BLAST(https://www.ncbi.nlm.nih.gov/ tools/primer-blast/)在线工具对设计的引物进行特异性检验[28]。引物委托上海英潍捷基生物技术有限公司合成。
1.4 多重PCR反应体系的建立
由于多重PCR使用的是大连宝生物工程有限公司生产的2×Multiplex PCR试剂盒(V2.0),因此,重点对影响多重PCR反应体系的引物退火温度以及扩增程序中延伸时间和循环数等重要参数分别进行单因素优化,其余试剂用量和扩增条件严格按照试剂盒推荐剂量和扩增条件。玉米大斑病菌和小斑病菌交配型的多重PCR检测特异引物的退火温度分别设置为62.0、61.6、60.7、59.1、57.2、55.6、54.5、54.0℃和56.0、55.6、54.7、53.1、51.2、49.6、48.5、48.0℃;多重PCR反应体系的延伸时间分别设置为30、45和60 s;循环数分别设为30、35和40次。扩增反应在Bio-Rad C1000TM型梯度PCR仪中进行。扩增结束后取7 µL扩增产物于120 v恒定电压下用含EB的1.5%琼脂糖电泳20 min,UVP BioDoc-ItTM凝胶成像系统检测并拍照。
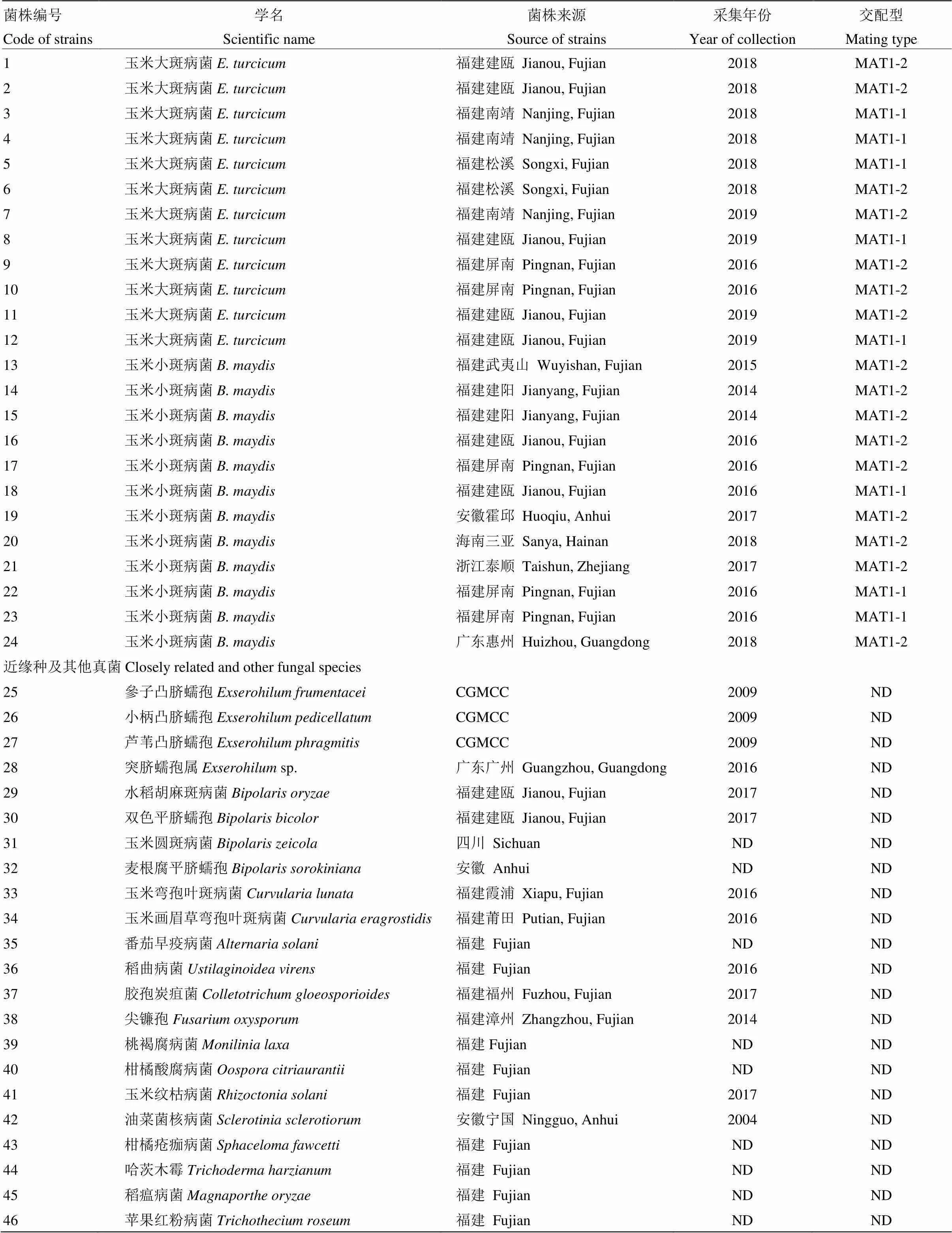
表1 供试菌株信息
CGMCC:中国普通微生物菌种保藏管理中心China General Microbiological Culture Collection Center;ND:不确定not determined
1.5 交配型多重PCR检测的验证
分别以12株玉米大斑病菌和小斑病菌交配型已知菌株的基因组DNA为模板(表1),利用优化的多重PCR反应体系进行扩增,以不加模板DNA为空白对照。每菌株5个重复,试验重复3次。检验设计的玉米大斑病菌和小斑病菌交配型多重PCR检测的准确性。
1.6 多重PCR灵敏度检测
取交配型为MAT1-1和MAT1-2型玉米大斑病菌菌株的基因组DNA各1份,将DNA模板按10倍梯度稀释,获得浓度梯度为100、10、1、10-1、10-2、10-3和10-4ng·μL-1;玉米小斑病菌MAT1-1和MAT1-2型菌株的DNA浓度梯度设置为10、1、10-1、10-2、10-3、10-4和10-5ng·μL-1,利用优化的多重PCR反应体系进行扩增,每25 μL PCR体系取上述各浓度DNA模板1 μL,以不加模板DNA为空白对照。每浓度梯度5个重复,试验重复3次。检验玉米大斑病菌和小斑病菌交配型多重PCR检测的灵敏度。
1.7 多重PCR特异性检测
选取玉米大斑病菌和小斑病菌MAT1-1和MAT1-2型菌株的基因组DNA各3份,分别以和近缘种以及其他14个属(种)真菌基因组DNA为阳性对照(表1),以不加模板DNA为空白对照。每菌株设置5个重复,试验重复3次。检验玉米大斑病菌和小斑病菌交配型多重PCR检测的特异性。
1.8 多重PCR检测体系的应用
从福建、安徽、海南、浙江和广东省采集田间感染玉米大斑病和小斑病的病叶标本,采用单孢分离纯化法获得田间玉米大斑病菌和小斑病菌菌株分别为129和194株[29-30],按上述方法提取各菌株的基因组DNA,利用本研究设计的交配型特异性引物和优化的多重PCR反应体系进行玉米大斑病菌和小斑病菌交配型检测。分别从福建省不同地区以及安徽、海南、浙江和广东省随机选取玉米大斑病菌或小斑病菌菌株5—10株。分别参考郭丽媛等[9]和OIDE等[31]方法与交配型已知的标准菌株进行室内杂交,以验证该多重PCR检测结果的准确性。
2 结果
2.1 多重PCR特异性引物的设计
通过DNAMAN 5.0软件的多重序列比对,利用Primer Premier 5.0软件筛选出4对较为理想的多重PCR引物用于玉米大斑病菌和小斑病菌交配型的检测。引物StMAT01-2F/R和StMAT02-3F/R分别用于检测玉米大斑病菌MAT1-1和MAT1-2交配型,预期PCR产物大小分别为816和132 bp;引物ChMAT01- 3F/R和ChMAT02-2F/R分别用于检测玉米小斑病菌MAT1-1和MAT1-2交配型,预期产物大小分别为490和136 bp(图1、表2)。Primer-BLAST检验结果表明,这4对引物的特异性较好,可用于后续试验。
2.2 多重PCR反应体系的建立
通过对引物的退火温度以及扩增程序中延伸时间和循环数的单因素优化,最终明确玉米大斑病菌交配型多重PCR检测体系(25 μL)为2×Multiplex PCR Mix 12.5 μL,引物各10 pmol,DNA模板100 ng,补充ddH2O至25 μL。反应程序:94℃ 5 min;94℃ 45 s,57.2℃ 45 s,72℃ 60 s,35个循环;最后72℃延伸10 min(表2)。
玉米小斑病菌交配型多重PCR检测体系(25 μL)为2×Multiplex PCR Mix 12.5 μL,引物各10 pmol,DNA模板100 ng,补充ddH2O至25 μL。反应程序:94℃ 5 min;94℃ 45 s,55℃ 45 s,72℃ 45 s,35个循环;最后72℃延伸10 min(表2)。
2.3 交配型多重PCR检测的验证
利用本研究设计的交配型特异引物和优化的反应体系分别对12株已知交配型的玉米大斑病菌和小斑病菌进行多重PCR检测,结果表明引物StMAT01- 2F/R对所有MAT1-1型的玉米大斑病菌菌株可扩增出一条816 bp的特异性条带,对交配型为MAT1-2型的菌株则未扩增出条带;相反,引物StMAT02-3F/R对所有MAT1-2型的玉米大斑病菌菌株可扩增出一条132 bp的特异性条带,而对MAT1-1型的菌株则未扩增出任何条带(图2-A)。引物ChMAT01-3F/R和ChMAT02-2F/R分别仅对交配型为MAT1-1和MAT1-2的玉米小斑病菌菌株扩增出一条490和136 bp的特异性条带(图2-B)。玉米大斑病菌和小斑病菌交配型的多重PCR检测结果与菌株的交配型完全一致,表明设计的交配型特异引物和建立的多重PCR反应体系准确性高,可用于玉米大斑病菌和小斑病菌交配型的快速检测。

红色和蓝色的方框及箭头分别表示玉米大斑病菌(A)和小斑病菌(B)交配型MAT1-1和MAT1-2型特异引物结合位点
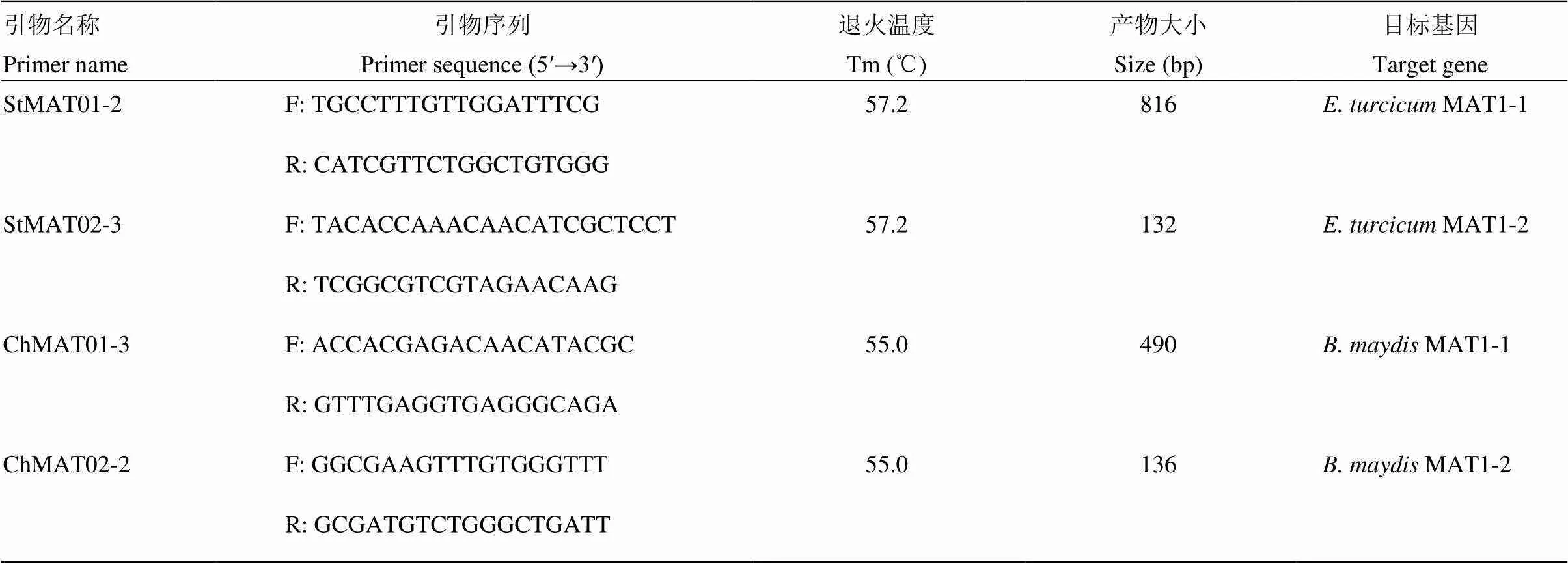
表2 玉米大斑病菌和小斑病菌交配型多重PCR检测的特异引物
2.4 多重PCR灵敏度检测
优化的多重PCR检测体系对玉米大斑病菌MAT1-1交配型的检测灵敏度为0.1 ng基因组DNA,而对MAT1-2交配型的检测灵敏度达0.01 ng基因组DNA(图3-A)。对玉米小斑病菌MAT1-1和MAT1-2交配型的检测灵敏度均为0.1 ng基因组DNA(图3-B)。
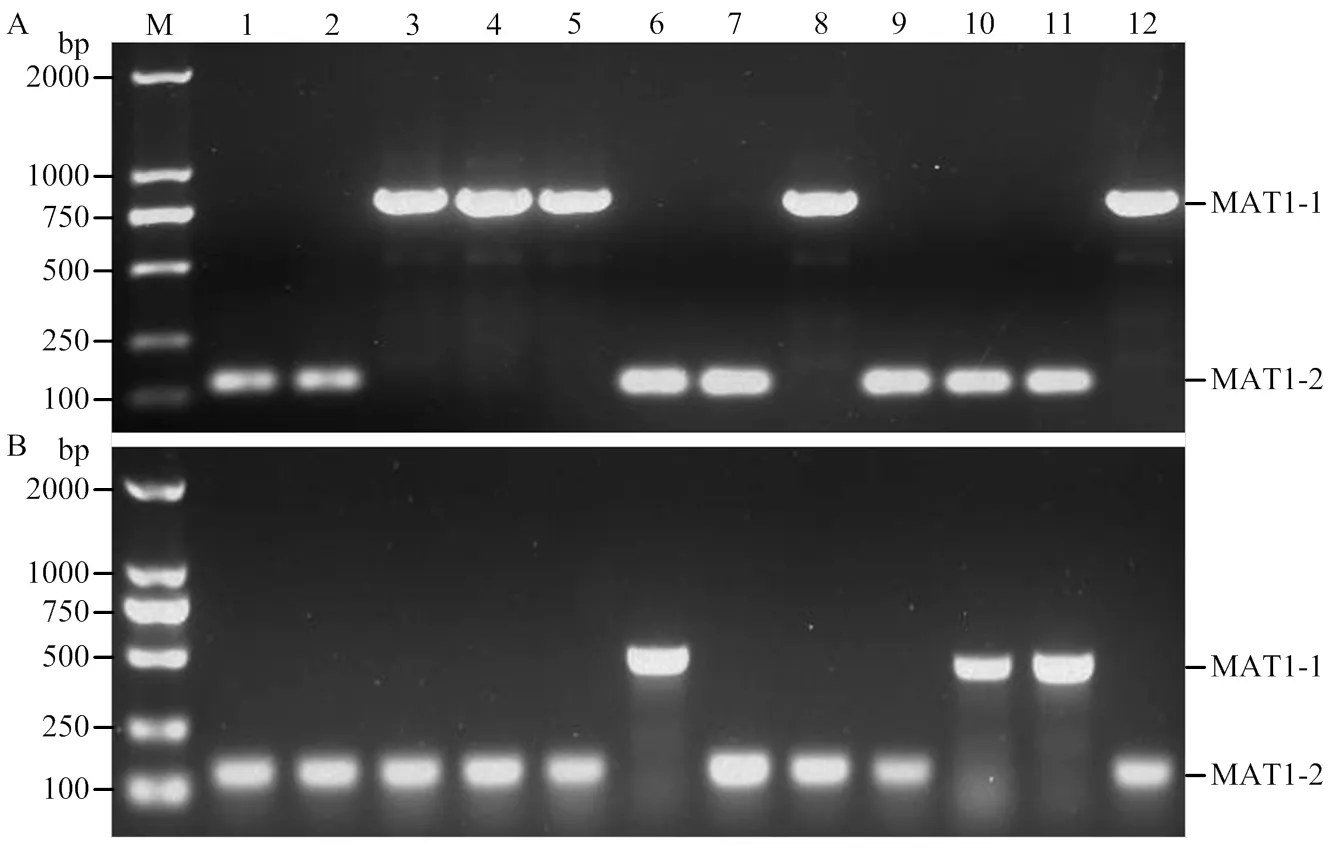
M:2 kb DNA ladder;1—12:表1中列举的12株交配型已知的玉米大斑病菌(A)和小斑病菌(B)菌株
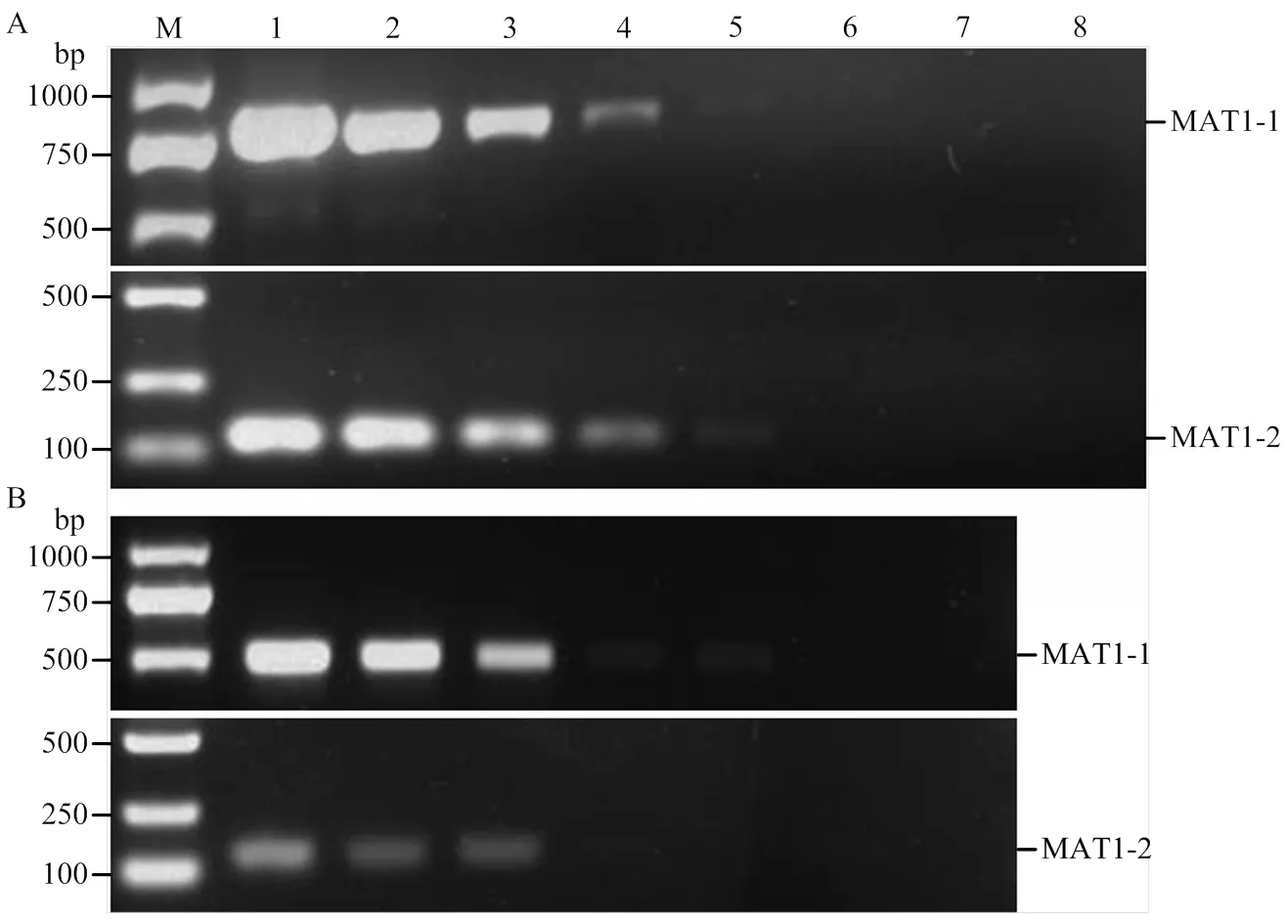
M:2 kb DNA ladder;1—7:(A)玉米大斑病菌MAT1-1和MAT1-2交配型菌株的基因组DNA的量为100、10、1、0.1、0.01、0.001和0.0001 ng Genomic DNA from MAT1-1 and MAT1-2 strains of E. turcicum at amounts of 100, 10, 1, 0.1, 0.01, 0.001 and 0.0001 ng, respectively;(B)玉米小斑病菌MAT1-1和MAT1-2交配型菌株的基因组DNA的量为10、1、0.1、0.01、0.001、0.0001和0.00001 ng Genomic DNA from MAT1-1 and MAT1-2 strains of B. maydis at amounts of 10, 1, 0.1, 0.01, 0.001, 0.0001 and 0.00001 ng, respectively;8:空白对照reagent only without DNA template
2.5 多重PCR特异性检测
建立的多重PCR检测体系中,引物StMAT01- 2F/R和StMAT02-3F/R分别仅对玉米大斑病菌MAT1-1和MAT1-2型菌株扩增出一条特异性条带,而对4株近缘种和14株其他真菌菌株未扩增出任何条带(图4-A)。引物ChMAT01-3F/R和ChMAT02- 2F/R分别仅对交配型为MAT1-1和MAT1-2的玉米小斑病菌扩增出一条490和136 bp的特异性条带,而对4株近缘种和14株其他真菌菌株未检测出特异性条带(图4-B)。

M:2 kb DNA ladder;1—6:分别为3株MAT1-1和MAT1-2的玉米大斑病菌(A)和小斑病菌(B)菌株Three MAT1-1 and MAT1-2 strains of E. turcicum (A) and B. maydis (B), respectively;7—10:表1所示的4株玉米大斑病菌(A)和小斑病菌(B)近缘种菌株Four strains of closely related species of Exserohilum (A) and Bipolaris (B), respectively, listed in Table 1;11—24:表1所示的14株其他真菌菌株Fourteen strains of other fungal species listed in Table 1
2.6 多重PCR检测体系的应用
分别从福建、安徽、浙江、广东和海南省分离的129和194株玉米大斑病菌和小斑病菌单孢菌株中提取基因组DNA,进行玉米大斑病菌和小斑病菌交配型的多重PCR检测。结果表明129和194株不同省份的玉米大斑病菌和小斑病菌均检测到MAT1-1和MAT1-2两种交配型,玉米大斑病菌和小斑病菌的交配型检测成功率均为100%(表3)。所有菌株均为无性态菌株,未检测到有性态菌株。室内杂交验证结果表明,与标准菌株杂交也能够成功检测出随机抽取菌株的交配型,且检测结果与多重PCR检测结果完全一致(表3)。上述结果表明,通过本研究设计的交配型特异引物和建立的多重PCR检测体系可对玉米大斑病菌和小斑病菌MAT1-1和MAT1-2两种交配型的菌株进行快速检测。
3 讨论
有性重组是植物病原菌遗传变异的重要来源,是病原菌强致病力新小种的出现、抗药性的产生以及高寄主适合度菌株形成的主要途径[32-33]。玉米大斑病和小斑病均为典型的气流传播性病害,可在田间短期内流行暴发,且该病原菌具有寄主适应性强、生理分化明显以及遗传变异性等特点[9,11,34]。因此,检测和监测玉米大斑病菌和小斑病菌的交配型,揭示其有性生殖和遗传变异机制,对玉米大斑病和小斑病的长期可持续控制具有重要意义。室内杂交是玉米大斑病菌和小斑病菌交配型的传统检测方法,该方法需要具备交配亲和性高和已知交配型的标准菌株,且从菌株分离纯化到获得理想的杂交检测结果耗时较长,不利于变异较快的玉米大斑病菌和小斑病菌大样本交配型的快速和及时检测。尽管GAFUR等建立了玉米小斑病菌交配型的PCR检测方法[35-36],但是,该方法或缺乏有效的灵敏性和特异性检验或操作步骤较为繁琐。因此,本研究分别建立了玉米大斑病菌和小斑病菌交配型的多重PCR检测方法,该方法具有更加快速、灵敏、准确、特异、可靠以及操作简便等特点,适合大样本交配型的快速、准确检测。
多重PCR是在一个反应体系里加入2对及以上引物进行PCR扩增,反应体系较为复杂,受诸多内外因素的影响[37]。特异引物设计的成功与否直接决定了多重PCR检测的成败。本研究利用已报道的玉米大斑病菌和小斑病菌交配型基因进行序列多重比对,在各保守区设计和筛选出错配率低、退火温度一致、扩增产物大小合适的4对交配型检测特异引物,引物经NCBI网站的Primer-BLAST在线工具进行特异性验证,最终分别成功建立了玉米大斑病菌和小斑病菌交配型的多重PCR检测方法。优化多重PCR反应体系和扩增条件是获得理想扩增结果的前提。本研究针对影响多重PCR扩增结果的重要参数,如引物的退火温度、扩增的延伸时间以及扩增循环数等分别进行单因素3—8个水平的系统优化,结果表明,引物的退火温度、扩增的延伸时间以及扩增循环数均对多重PCR反应体系有明显影响。其中特异引物的退火温度是决定实验成败的关键因素,一般退火温度越低,扩增的条带模糊、特异性低;退火温度越高,不利于大片段的扩增。因此,本研究通过梯度PCR扩增优化出玉米大斑病菌和小斑病菌交配型多重PCR检测引物的最佳退火温度分别为57.2℃和55.0℃。尽管本研究未对多重PCR反应体系中Taq聚合酶用量、dNTPs浓度、Mg2+浓度以及模板DNA浓度进行系统优化,但是根据预混液的推荐剂量进行多重PCR扩增也能获得理想的结果,表明本研究建立的玉米大斑病菌和小斑病菌交配型多重PCR检测方法的适用性较好。
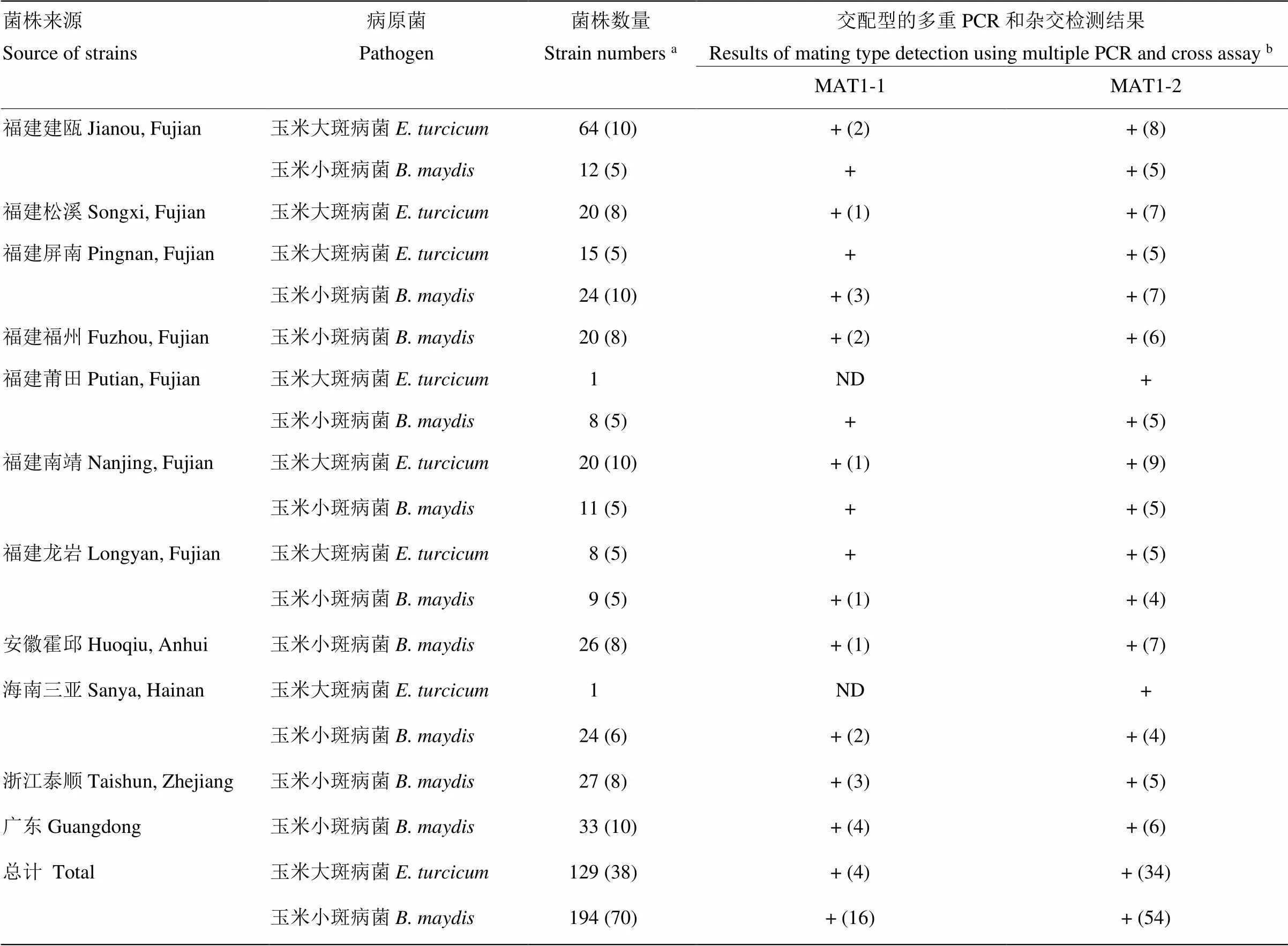
表3 不同地理来源的玉米大斑病菌和小斑病菌交配型的多重PCR和杂交检测
a括号内数字表示随机抽取用于杂交实验的菌株数The number in parentheses represents the number of strains which were randomly selected for cross assays;b“+”:多重PCR方法成功检测出所有待测菌株的交配型Mating types of all tested strains were successfully detected using multiple PCR method;小括号内数字表示杂交检测出的菌株数,且与多重PCR检测结果完全一致The number in parentheses indicates the number of strains detected by cross assays, which are completely consistent with the results of multiple PCR detection;ND:未检测No detection
理想的交配型PCR检测方法应具有较高的灵敏度和特异性[36]。多重PCR的灵敏度越高,模板DNA量的需求就越低。本研究建立的玉米大斑病菌和小斑病菌交配型多重PCR检测最低灵敏度为0.1 ng基因组DNA,该剂量的模板浓度极易从病原菌的菌丝或分生孢子中通过DNA小剂量提取法获得[38],从而降低了对模板DNA的要求,提高了多重PCR交配型检测的成功率。引物的特异性是多重PCR的另一重要因素[36],本研究建立的玉米大斑病菌和小斑病菌交配型多重PCR检测方法不仅能够很好地区分玉米大斑病菌和小斑病菌的交配型,而且能够分别从其相应的和近缘种和其他14个属种间区分开来,避免了由于近缘种干扰而产生的假阳性,提高了多重PCR交配型检测的准确性。此外,由于本研究建立的玉米大斑病菌和小斑病菌交配型多重PCR检测方法具有较高的灵敏度和特异性,能够分别将其从相应的近缘种和其他属种区分来(图4),因此,该多重PCR检测体系还可以用于玉米大斑病和小斑病的田间早期快速诊断和病害的预测预报,为有效预防和控制玉米大斑病和小斑病,保障玉米安全优质生产提供了技术支持。
通过建立的玉米大斑病菌和小斑病菌交配型多重PCR检测方法分别对来自福建、安徽、浙江、广东和海南省的129和194株玉米大斑病菌和小斑病菌的交配型进行检测,结果显示两种病原菌的交配型检测成功率均为100%,且多重PCR检测结果与杂交验证结果吻合,表明该检测方法准确性高、可靠性好。通过交配型检测发现,不同省份或地区的玉米大斑病菌和小斑病菌均同时存在MAT1-1和MAT1-2两种交配型,所有菌株均为无性态菌株,未检测到有性态菌株。该研究结果与郭建国等均检测到玉米大斑病菌田间有性态菌株的结果不一致[12,39-40]。本研究未检测到有性态菌株,说明上述采集地田间菌株发生有性生殖的风险性较低。但是,不同交配型菌株的同时存在,增加了有性重组的可能性,暗示未来病菌群体的遗传结构和遗传多样性可能发生更加复杂的变化。因此,将来工作的重点应该是加强对玉米大斑病菌和小斑病菌田间交配型、群体遗传结构和遗传多样性的监测以及病原菌遗传变异机制的深入研究。
4 结论
通过设计交配型检测特异引物并对反应条件进行优化,分别建立了玉米大斑病菌和小斑病菌交配型的多重PCR检测方法。该方法具有灵敏度高、特异性好和操作简便的特点,能够准确、快速地检测出玉米大斑病菌和小斑病菌MAT1-1和MAT1-2两种交配型,为玉米大斑病菌和小斑病菌交配型的田间分布和监测以及有性遗传研究提供了可靠的技术方法。
[1] WEEMS J D, BRADLEY C A.race population distribution in the north central United States., 2018, 102(2): 292-299.
[2] DAI Y L, GAN L, RUAN H C, SHI N N, DU Y X, LIAO L, WEI Z X, TENG Z Y, CHEN F R, YANG X J. Sensitivity ofto three demethylation inhibitor fungicides, propiconazole, diniconazole and prochloraz, and their efficacy against southern corn leaf blight in Fujian Province, China., 2018, 152(2): 447-459.
[3] WANG M, WANG S Q, MA J, YU C J, GAO J X, CHEN J. Detection ofraces in South China., 2017, 165(10): 681-691.
[4] HUMAN M P, BARNES I, CRAVEN M, CRAMPTON B G. Lack of population structure and mixed reproduction modes infrom South Africa., 2016, 106(11): 1386-1392.
[5] 代玉立, 甘林, 阮宏椿, 石妞妞, 杜宜新, 陈福如, 杨秀娟. 福建省丙环唑不同敏感性玉米小斑病菌的遗传多样性和致病性. 植物病理学报, 2019, 49(1): 64-74.
DAI Y L, GAN L, RUAN H C, SHI N N, DU Y X, CHEN F R, YANG X J. Genetic diversity and pathogenicity of different propiconazole-sensitive isolates ofin Fujian Province., 2019, 49(1): 64-74. (in Chinese)
[6] 赵书文, 杨秀林, 郭东. 玉米大斑病的流行原因与综合治理措施. 中国植保导刊, 2005, 25(3): 10-12.
ZHAO S W, YANG X L, GUO D. Epidemic causes ofand its integrated management measures., 2005, 25(3): 10-12. (in Chinese)
[7] 裴静宇, 高锋, 杨彦文, 张姼, 杨信东. 玉米小斑病重要流行环节的初步定量研究I. 孢子萌发侵入、病斑潜育显症及扩展. 吉林农业大学学报, 2007, 29(1): 28-32.
PEI J Y, GAO F, YANG Y W, ZHANG S, YANG X D. Preliminary quantitative studies on important epidemic links ofⅠ. The spores germination penetration, incubation and expand of lesion., 2007, 29(1): 28-32. (in Chinese)
[8] 陈利锋, 徐敬友. 农业植物病理学. 3版. 北京: 中国农业出版社, 2007.
CHEN L F, XU J Y... Beijing: China Agriculture Press, 2007. (in Chinese)
[9] 郭丽媛, 贾慧, 曹志艳, 谷守芹, 孙淑琴, 董金皋. 玉米大斑病菌有性杂交后代的交配型与寄生适合度分化. 中国农业科学, 2013, 46(19): 4058-4065.
GUO L Y, JIA H, CAO Z Y, GU S Q, SUN S Q, DONG J G. Analysis on mating type and parasitic fitness diversity in sexual hybridization offsprings of., 2013, 46(19): 4058-4065. (in Chinese)
[10] 王玉萍, 王晓鸣, 马青. 我国玉米大斑病菌生理小种组成变异研究. 玉米科学, 2007, 15(2): 123-126.
WANG Y P, WANG X M, MA Q. Races of, causal agent of northern leaf blight in China., 2007, 15(2): 123-126. (in Chinese)
[11] 冯为民. 黄淮海地区玉米小斑病菌变异研究[D]. 保定: 河北大学, 2012.
FENG W M. The study of differentiation ofin Huang-Huai-Hai region[D]. Baoding: Hebei University, 2012.(in Chinese)
[12] 郭建国, 杨凤珍, 杜蕙, 金社林, 张新瑞. 甘肃省玉米大斑病菌生理小种结构组成与交配型测定. 植物保护, 2018, 44(6): 85-90.
GUO J G, YANG F Z, DU H, JIN S L, ZHANG X R. Structural composition of physiological races and mating types ofin Gansu., 2018, 44(6): 85-90. (in Chinese)
[13] CHAO C C T, ELLINGBOE A H. Selection for mating competence inpathogenic to rice., 1991, 69(10): 2130-2134.
[14] NOTTEGHEM J L, SILUÉ D. Distribution of the mating type inpopulations pathogenic on rice., 1992, 82(4): 421-424.
[15] CHÉRIF M, CHILVERS M I, AKAMATSU H, PEEVER T L, KAISER W J. Cloning of the mating type locus from(teleomorph:) and development of a multiplex PCR mating assay forspecies., 2006, 50(3): 203-215.
[16] 史文琦, 龚双军, 曾凡松, 薛敏峰, 杨立军, 喻大昭. 小麦白粉病菌有性生殖与自然群体交配型检测. 植物病理学报, 2016, 46(5): 645-652.
SHI W Q, GONG S J, ZENG F S, XUE M F, YANG L J, YU D Z. Sexual reproduction and detection of mating-type off. sp.populations., 2016, 46(5): 645-652. (in Chinese)
[17] BREWER M T, CADLE-DAVIDSON L, CORTESI P, SPANU P D, MILGROOM M G. Identification and structure of the mating-type locus and development of PCR-based markers for mating type in powdery mildew fungi., 2011, 48(7): 704-713.
[18] TAKAN J P, CHIPILI J, MUTHUMEENAKSHI S, TALBOT N J, MANYASA E O, BANDYOPADHYAY R, SERE Y, NUTSUGAH S K, TALHINHAS P, HOSSAIN M, BROWN A E, SREENIVASAPRASAD S.populations adapted to finger millet and rice exhibit distinctive patterns of genetic diversity, sexuality and host interaction., 2012, 50(2): 145-158.
[19] DE MICCOLIS ANGELINI R M, ROTOLO C, POLLASTRO S, FARETRA F. Molecular analysis of the mating type () locus in strains of the heterothallic ascomycete., 2016, 65(8): 1321-1332.
[20] FOSTER S J, SINGH G, FITT B D L, ASHBY A M. Development of PCR based diagnostic techniques for the two mating types of(light leaf spot) on winter oilseed rape (ssp.)., 1999, 55(2): 111-119.
[21] LINDE C C, ZALA M, CECCARELLI S, MCDONALD B A. Further evidence for sexual reproduction inbased on distribution and frequency of mating-type alleles., 2003, 40(2): 115-125.
[22] ALLIOUI N, SIAH A, BRINIS L, REIGNAULT P, HALAMA P. Mating type distribution provides evidence for sexual reproduction ofin Algeria., 2014, 36(4): 475-481.
[23] PEARCE T L, SCOTT J B, HAY F S, PETHYBRIDGE S J. Mating-type gene structure and spatial distribution ofin pyrethrum fields., 2016, 106(12): 1521-1529.
[24] TURGEON B G, YODER O C. Proposed nomenclature for mating type genes of filamentous ascomycetes., 2000, 31(1): 1-5.
[25] RAMATHANI I, BIRUMA M, MARTIN T, DIXELIUS C, OKORI P. Disease severity, incidence and races ofon sorghum in Uganda., 2011, 131(3): 383-392.
[26] TURGEON B G, BOHLMANN H, CIUFFETTI L M, CHRISTIANSEN S K, YANG G, SCHÄFER W, YODER O C. Cloning and analysis of the mating type genes from., 1993, 238(1/2): 270-284.
[27] 代玉立, 甘林, 阮宏椿, 石妞妞, 杜宜新, 陈福如, 杨秀娟. 福建省鲜食玉米小型叶斑病的病原菌鉴定. 福建农业学报, 2017, 32(12): 1341-1349.
DAI Y L, GAN L, RUAN H C, SHI N N, DU Y X, CHEN F R, YANG X J. Pathogen identification of small leaf spots on sweet corn plants in Fujian., 2017, 32(12): 1341-1349. (in Chinese)
[28] YE J, COULOURIS G, ZARETSKAYA I, CUTCUTACHE I, ROZEN S, MADDEN T L. Primer-blast: a tool to design target-specific primers for polymerase chain reaction., 2012, 13: 134.
[29] 高金欣, 高增贵, 张小飞, 张硕, 陈璐. 一种简捷的玉米大斑病病菌单孢分离方法. 微生物学通报, 2010, 37(10): 1548-1550.
GAO J X, GAO Z G, ZHANG X F, ZHANG S, CHEN L. A simple and feasible method for single-spore isolation of., 2010, 37(10): 1548-1550. (in Chinese)
[30] DAI Y L, YANG X J, GAN L, CHEN F R, RUAN H C, DU Y X, SHI N N, GAO Z M. First report of southern leaf blight caused byon corn (L.) in Fujian Province, China., 2016, 100(8): 1781.
[31] OIDE S, KRASNOFF S B, GIBSON D M, TURGEON B G. Intracellular siderophores are essential for ascomycete sexual development in heterothallicand homothallic., 2007, 6(8): 1339-1353.
[32] GLASS L N, KULDAU G A. Mating type and vegetative incompatibility in filamentous ascomycetes., 1992, 30: 201-224.
[33] BI Y, HU J, CUI X, SHAO J, LU X, MENG Q, LIU X. Sexual reproduction increases the possibility thatwill develop resistance to dimethomorph in China., 2014, 63(6): 1365-1373.
[34] JINDAL K K, TENUTA A U, WOLDEMARIAM T, ZHU X Y, HOOKER D C, REID L M. Occurrence and distribution of physiological races ofin Ontario, Canada., 2019, 103(7): 1450-1457.
[35] GAFUR A, TANAKA C, OUCHI S, TSUDA M. A PCR-based method for mating type determination in., 1997, 38(4): 455-458.
[36] DAI Y L, GAN L, RUAN H C, SHI N N, DU Y X, CHEN F R, YANG X J. A PCR method to detect mating types of., 2018, 40(3): 358-367.
[37] 刘芮池, 程有普, 柴阿丽, 石延霞, 谢学文, 帕提古丽, 李宝聚. 蔬菜土传病原菌三重PCR检测体系的建立与应用. 中国农业科学, 2019, 52(12): 2069-2078.
LIU R C, CHENG Y P, CHAI A L, SHI Y X, XIE X W, PATIGULI, LI B J. Establishment and application of a triplex PCR detection system for vegetable soil-borne pathogens., 2019, 52(12): 2069-2078. (in Chinese)
[38] LU S W, PLATZ G J, EDWARDS M C, FRIESEN T L. Mating type locus-specific polymerase chain reaction markers for differentiation off.andf., the causal agents of barley net blotch., 2010, 100(12): 1298-1306.
[39] 孙淑琴, 温雷蕾, 董金皋. 玉米大斑病菌的生理小种及交配型测定. 玉米科学, 2005, 13(4): 112-113, 123.
SUN S Q, WEN L L, DONG J G. Identification of physiological races and mating type of., 2005, 13(4): 112-113, 123. (in Chinese)
[40] 王利智, 康志钰, 吴景芝, 吴毅歆, 毛自朝, 何月秋. 云南省玉米大斑病菌生理小种的鉴定及交配型分析. 华中农业大学学报, 2011, 30(2): 187-192.
WANG L Z, KANG Z Y, WU J Z, WU Y X, MAO Z C, HE Y Q. Identification of physiological races and analysis of mating type ofin Yunnan Province., 2011, 30(2): 187-192. (in Chinese)
Establishment and Application of a Multiple PCR Method to Detect Mating Types ofand
DAI YuLi1, GAN Lin1, TENG ZhenYong2, YANG JingMin3, QI YueYue4, SHI NiuNiu1, CHEN FuRu1, YANG XiuJuan1
(1Institute of Plant Protection, Fujian Academy of Agricultural Sciences/Fujian Key Laboratory for Monitoring and Integrated Management of Crop Pests, Fuzhou 350013;2Fujian Seed Management Station, Fuzhou 350001;3Jianou Municipal Bureau of Agriculture and Rural Affairs, Jianou 353100, Fujian;4Zhejiang Tianfeng Biological Science Co. Ltd, Jinhua 321000, Zhejiang)
【Objective】Northern corn leaf blight (NCLB) and southern corn leaf blight (SCLB), caused byand, respectively, are the most important foliar fungal diseases affecting maize production. The objective of this study is to establish a multiple PCR method to detect mating types ofand, and to provide a technical method for the study of mating type distribution in the field and sexual reproduction ofand.【Method】Mating type-specific primers for the two pathogens were designed on the basis of mating type gene sequences of(accession numbers: GU997138 for MAT1-1; GU997137 for MAT1-2) and(accession numbers: X68399 for MAT1-1; X68398 for MAT1-2) obtained from GenBank, and the important parameters of primer annealing temperatures, extension times and amplification cycles in the amplification program were optimized using the single factor method. A multiple PCR method was established to detect mating types ofand, and the sensitivity and specificity of the multiple PCR were also assessed. Meanwhile, the mating types of 129 strains ofand 194 strains offrom field-collections were detected by the multiple PCR to determine the adaptability of the established method.【Result】The expected 816, 132 bp (), and 490, 136 bp () target fragments were amplified specifically using the multiple PCR with mating type-specific primers of StMAT01-2F/R, StMAT02-3F/R, and ChMAT01-3F/R, ChMAT02-2F/R from MAT1-1 and MAT1-2 strains, respectively. A 25 μL PCR reaction system consisted of 12.5 μL 2×Multiplex PCR Mix, 10 pmol each primer, and 100 ng DNA template. The annealing temperatures forandwere 57.2℃ and 55.0℃, respectively, and the number of amplification cycles was 35. The multiple PCR method could reliably detect mating types ofat 0.1 ng genomic DNA for MAT1-1 or 0.01 ng DNA for MAT1-2 from single-spore strains, while the sensitivity of the multiple PCR to detect mating types ofwas 0.1 ng genomic DNA for both MAT1-1 and MAT1-2 from pure culture strains. The method exhibited specificity in differentiating mating types ofandfrom their closely-related species, as well as 14 other fungal genera. The results of mating types detection ofandstrains from different geographical origins indicated that the multiple PCR could reliably detect mating types of 129 and 194 strains ofand, respectively. These results were consistent with the verification results of laboratorial cross assays with random selected strains from different locations.【Conclusion】The established multiple PCR method for mating types detection ofandin this study was characterized as high sensitivity, specificity and user-friendly control, it could accurately and rapidly detect mating types ofand. This study provides a reliable technique and approach for the study of distribution and monitoring of mating types in the field and sexual reproduction ofand.
;; mating type; multiple PCR; sensitivity; specificity
2019-08-13;
2019-09-18
福建省属公益类科研院所专项(2017R1025-3,2018R1025-1)、国家重点研发计划(2018YFD0200706)、福建省农业科学院青年自由探索项目(AA2018-14)、福建省农业科学院青年科技英才百人计划(YC2016-4)、福建省农业科学院植物保护创新团队(STIT2017-1-8)
代玉立,E-mail:dai841225@126.com。通信作者杨秀娟,E-mail:yxjzb@126.com
(责任编辑 岳梅)
