柑橘不同类型砧木的种子和苗期性状
2020-02-25朱世平王福生陈娇余歆余洪罗国涛胡洲冯锦英赵晓春洪棋斌
朱世平,王福生,陈娇,余歆,余洪,罗国涛,胡洲,冯锦英,赵晓春,洪棋斌
柑橘不同类型砧木的种子和苗期性状
朱世平1,王福生1,陈娇2, 3,余歆1,余洪1,罗国涛1,胡洲1,冯锦英1,赵晓春1,洪棋斌1
(1西南大学/中国农业科学院柑桔研究所/国家柑桔工程技术研究中心,重庆 400712;2西南大学园艺园林学院,重庆 400716;3四川省富顺县农业农村局,四川富顺 643200)
【目的】优良的砧木能促进果树生长、提高果实产量和品质、增强植株的抗逆性和适应性。目前我国柑橘生产上使用的砧木品系混杂,生长参差不齐,抗逆性差异明显,严重影响了苗木质量。通过对各种种质类型柑橘砧木的连续多年评价,分析不同遗传种质砧木的种子和苗期特征,鉴定影响砧木苗木质量的关键性状,建立规范柑橘砧木的评价标准,为筛选优良砧木单系提供指导。【方法】以104份柑橘砧木种质为材料,连续5年评价单果种子数、种子饱满度、千粒重和胚型等种子性状及播种后的出苗率、黄化率、立枯率、株高和茎粗等苗期性状,并对这些指标进行相关性、主成分分析以及不同年份间的变异系数分析。利用保守的直系同源序列(conserved ortholog sequences,COS)分子标记技术评价部分不同胚型砧木种质幼苗遗传背景的一致性,分析部分种质中微型反向重复转座元件(miniature inverted-repeat transposable element,MITE)片段插入与胚型的关联性。【结果】(1)枳、枳杂种和香橙的果实为多核和较多核,种子为混胚和多胚,种子饱满,大部分材料的种子千粒重为200 g以上;而宽皮柑橘为少核、较多核和多核,多胚,种子中等饱满,约一半种质的种子千粒重为100 g以下。(2)在苗期特性方面,香橙出苗率最高,黄化率较低,其幼苗对立枯病比较敏感;枳及其杂种的出苗率较高,黄化率也较高,但耐立枯病;宽皮柑橘的出苗率则较低,幼苗发生黄化和立枯的比例也比较低。比较播种后10个月内的幼苗生长势,枳的生长势最强,其次是枳杂种和香橙,而宽皮柑橘的生长势最弱。(3)对种子和苗期性状指标的相互关系及主成分分析结果表明,除黄化率和立枯率外,单果种子数、千粒重、饱满度、单胚比例、多胚比例、胚型、出苗率、幼苗株高和茎粗等指标之间具有极显著(<0.01)或显著(<0.05)的相关性,且不同指标对主成分的方差贡献率有明显差异。通过主成分分析能够清楚地区分枳、枳杂种、香橙、宽皮柑橘及其他砧木的种质类型。除黄化率外,种子性状和苗期表现的指标在不同年份间的变异系数相对较低,说明这些性状较为稳定。(4)COS分子标记结果显示,单胚宽皮柑橘幼苗的变异程度比单胚枳及其他多胚或混胚类型种质的变异程度高;在单胚的柚和宽皮柑橘等种质中没有MITE片段的插入,而在多胚的香橙、宽皮柑橘、枳杂种等种质中有插入,但是在枳种质中无论是单胚还是多胚均没有检测到MITE片段的插入。【结论】柑橘砧木种子性状和幼苗性状指标之间高度相关,通常种子性状优异的砧木种质,其幼苗质量也较好。枳的种子和幼苗(播种后10个月)综合性状最好,其次是枳杂种和香橙,宽皮柑橘的种子和苗期综合性状最差。
柑橘砧木;种子性状;胚型;出苗率;一致性
0 引言
【研究意义】嫁接是现代园艺植物生产中应用最广的一种繁殖技术,在中国最早被用于柑橘生产[1]。砧木是嫁接的最基本要素之一。优良砧木能促进果树早结丰产,提高果实品质,增强抗病性和对环境的适应能力[2]。美国、巴西等发达国家在20世纪30年代就开始标准化工厂育苗,培育健壮整齐的无病毒容器苗。我国在20世纪90年代开始标准化、规模化,培育柑橘无病毒容器苗,并从国外引进了柑橘砧木新品种如‘卡里佐’枳橙、‘施文格’枳柚等。但由于受柑橘砧木研究滞后和商业化砧木采种园不规范等因素的限制,目前只有个别砧木品种是采用纯系种子育苗,如‘卡里佐’枳橙。对大部分柑橘砧木而言,生产上使用的种子均是同一类型的混合品系种子,而不是单一品种(系)的种子,使用上也没有进行分级处理,导致在育苗过程中出现砧木的出苗率不稳定,种苗生长势不一致,整齐度较差等问题。调查结果显示,很多苗圃因为砧木种子品种纯度不够或未分级,以及嫁接成活率低等造成每年约15%—35%的容器苗不能按时出圃,一方面增加了育苗成本,另一方面严重影响柑橘苗木的质量。因此,对柑橘砧木种质进行种子性状及苗期评价,筛选出苗率高、长势强、整齐度好的优良单系砧木对柑橘规模化、优质化苗木繁育意义重大。【前人研究进展】枳、香橙、枳橙、红橘和酸柚等是我国柑橘的主要砧木,其中枳的应用历史最久,范围最广,面积最大,数量最多。在长期的进化和选择过程中,出现许多不同的株系变异,而且相同类型砧木有遗传多样性丰富的不同种质[3-5]。但在实际生产中通常没有区分这些变异株系,而是混合使用。万良珍和周开隆[6]以9份枳单系为砧木,开展了‘哈姆林’甜橙的砧木比较试验,发现枳的不同单系对‘哈姆林’的生长、产量和果实品质的影响有显著差异。在生产上,枳砧具有许多优良特性,如早结丰产性好,果实品质优,抗衰退病、抗流胶病、抗根结实,线虫病,抗寒能力强等,但同时也存在一些缺点,如不耐盐,在碱性条件下容易缺铁,对柑橘裂皮病、碎叶病等敏感,在挂果较多的情况下树体易早衰等。戴胜根等[7]发现不同枳单系对嫁接甜橙后的黄化有不同程度的影响。以上结果说明,相同类型砧木的不同单系间存在较为明显的遗传差异,这些差异对嫁接亲和性及产量、品质等都有影响。因此,需要加强不同砧木种质的系统评价,以筛选更加优良的单系砧木。砧木通常用种子进行实生繁殖。实生繁殖的砧木苗根系发达,生长健壮且容易培育[8]。另外,与营养繁殖方式相比,种子不易携带病毒等病原,能保证苗木健康。目前,对柑橘砧木资源种子特性的研究报道较少,砧木的苗期评价也多侧重于抗逆性方面,如耐旱性[9-11]和对pH的适应性[12-13]。解凯东等[14]比较了4个体细胞杂种砧木种质的种子特性,发现其中3个杂种组合的单果种子数较多,种子特性较优,适于作砧木应用。但是作为优良的柑橘砧木,必须同时具备优良的种子特性、苗期性状和抗逆性等,需要对多种性状进行综合评价和筛选。【本研究切入点】我国是世界柑橘的重要起源中心,砧木资源非常丰富。但是由于我国柑橘砧木研究起步较晚,研究滞后,目前尚未有人工培育的具有自主知识产权的柑橘砧木新品种报道,也未对我国砧木资源开展系统评价,更没有建立比较权威的柑橘砧木新品种评价的国家或行业标准。这导致柑橘砧木使用品系混杂,柑橘苗木质量参差不齐,严重影响我国柑橘产业的健康和可持续发展。【拟解决的关键问题】本研究以104份砧木资源为材料,对单果种子数、胚型等种子特性和苗期表现等影响柑橘苗木质量的相关指标进行连续5年的系统调查和评价,旨在筛选出优良的砧木单系用于生产,同时为建立我国柑橘砧木新品种筛选和评价标准提供参考和借鉴。
1 材料与方法
1.1 试验材料
2013—2017年连续5年采集104份砧木种质的成熟果实,包括枳、枳杂种、香橙、宽皮柑橘以及其他种类的砧木,收集种子进行评价。所有试验材料均采自中国农业科学院柑桔研究所国家果树种质(重庆)柑橘圃。对20份不同胚型的砧木种质进行与胚型紧密相关的微型反向重复转座元件(MITE)片段插入的检测[15-16]。利用保守的直系同源序列(COS)分子标记[17]对不同胚型的种质KPJ-023、KPJ-030()、Z-006、Z-022、Z-040()、ZZ-030(hybrid)、XC-011、XC-013()等材料进行幼苗的一致性检测。
1.2 试验方法
1.2.1 种子的收集、处理及相关参数的测定
(1)单果种子数的计算及种子处理:根据不同砧木种质本身的物候期和生物学特性确定采集时间,在果实达到生理成熟后采收至实验室进行种子相关指标的评价。枳的采集时间为9月下旬,枳杂种的采集时间为10—11月,其余砧木资源的采集时间为12月。每份砧木随机采集50个果实并取出种子,随机记录其中20个果实的单果种子数,然后计算每个果实的平均种子数。根据平均每个果实的种子数量,分为无核、少核、较多核和多核4个级别,即:1—2粒为无核,3—5粒为少核,6—20粒为较多核,20粒以上(含20粒)为多核[8]。种子取出后用0.5%的温热生石灰溶液处理2—3 min,以洗净表面包裹的果胶等黏稠物质。对于表面黏稠物质较多的枳及其杂种种子,需要重复处理几次。处理后用自来水冲洗干净,然后在(52± 1)℃热水中浸泡10 min进行热处理消毒,处理完成后立即放入自来水中使种子尽快降温,然后在通风较好的弱光条件下将种子晾干至种皮发白,表面无明显水汽后装入塑料袋备用[18]。
(2)种子饱满度调查和千粒重的测定:根据种子种皮的充实程度,将种子分为饱满、中等饱满和不饱满3个等级。1 000粒种子中有90%以上的种子为正常大小,种皮光滑没有皱褶,且正常条件下干燥后种皮紧密包被胚的为饱满;1 000粒种子中有80%以上的种子为正常大小,种皮有轻微皱褶且包被胚较为紧密的为中等饱满;1 000粒种子中90%以上的种子较小,种皮有皱褶且种皮包被胚比较松散的为不饱满。每份砧木种子随机选择100粒种子称重,计算千粒重,每份材料重复3次。
1.2.2 胚型分类 每份砧木随机挑选50粒种子,剥除种皮,在饱和湿度条件下静置12 h后,多胚会互相自然分开或胚与胚之间变得比较松散,然后用镊子将其完全分开,观察并记录胚的数量。每粒种子只有一个胚为单胚,有两个及以上胚的为多胚。根据单胚和多胚的种子数分别计算单胚比例和多胚比例。单胚比例≥94%则为单胚类型,10%≤单胚比例<10%为混胚类型,单胚比例<10%为多胚类型[19]。单(多)胚比例的计算如下:
单(多)胚比例=(单(多)胚种子数/50)×100%
1.2.3 种子的播种、苗期观察和统计 每年12月下旬至1月上旬播种。种子在播种前先用0.1%的高锰酸钾溶液浸泡30 min,然后在自来水中浸泡4—5 h,置于垫有湿润滤纸的培养皿中,28℃条件下暗培养3—5 d至萌发[20],最后播种到温室中的96孔育苗盘(2013—2016年)或栽培槽(2017年)。每份材料播种100粒种子,所用的基质为草炭﹕河沙﹕椰糠(体积比为1﹕1﹕1)。播种3个月后统计出苗数、黄化苗数、感染立枯病植株数,并按照以下公式计算出苗率、黄化率和立枯病感病率(立枯率)。
出苗率=出苗数/播种种子数×100%
黄化率=黄化苗数/出苗数×100%
立枯率=感病苗数/出苗数×100%
为比较不同砧木的生长势,2013—2016年在播种4—5个月后测定砧木幼苗的株高和茎粗,2017年在播种9个月后测定株高和茎粗。
1.2.4 不同胚型种质的MITE插入片段的检测 根据Wang等[15]和Shimada等[16]的报道,MITE在含有RWP结构域的基因启动子的插入是引起柑橘单胚和多胚性状分离的主要原因,因此,利用MITE特异正向引物5′-GTTACTTGGAGACGGCCTAACG-3′和反向引物5′-TCGATCATGTAATGCTGACTC-3′(Shimada等[16])检测不同胚型的20份砧木种质的MITE片段插入。采用Premix TaqTM (TaKaRa TaqTM Version 2.0)(宝日医生物技术(北京)有限公司,中国)酶扩增目的片段,Premix酶混合液为10 μL,引物各2 μL,2 μL DNA模板,补水至20 μL,置于Biometra PCR仪中进行扩增,扩增程序参照王炯等[17]方法,并部分修改为:96℃预变性4 min,94℃变性30 s,64℃退火40 s,72℃延伸40 s,在每个循环将退火温度降低0.5℃,循环10次;94℃变性30 s,58℃退火40 s,72℃延伸40 s,循环22次;72℃延伸5 min。取6 μL PCR产物用1.5%琼脂糖凝胶电泳检测。
1.2.5 砧木幼苗一致性的分子标记检测 在种子胚型鉴定的基础上,分别选择单胚、多胚和混胚3种胚型共8份种质(单胚:KPJ-023、Z-040;混胚:XC-011、ZZ-030、Z-022;多胚,XC-013、KPJ-030、Z-006),随机选取分布在全基因组中的27个COS分子标记(每条染色体上随机选3个标记),对砧木幼苗进行一致性鉴定。以采集种子的母树为对照,从种子播种后的幼苗中随机选取25个单株采集混合样品提取基因组DNA,每份种质分别取4份25个单株的混合样品,然后进行分析。如果4份混合样品的PCR产物电泳结果中有一个混合样品出现与母本不一样的带型,说明这个混合样品中至少有一个单株在这个位点发生了变异,即认为这100个单株中至少有1株发生了遗传变异;如果4个混合样中均出现了与母本不一样的带型,即认为这100个单株中至少有4株发生了变异,因此幼苗的变异株率(%)≥4%。位点变异率为发生变异的位点个数占27个位点的百分比,反映幼苗发生遗传变异的程度。27对COS marker的引物序列见王炯[21]。PCR反应体系为优化的15 μL体系:1×PCR Buffer,1.5 mmol·L-1的Mg2+,0.2 mmol·L-1的dNTPs,0.33 nmol·L-1的上、下游引物,1 U的DNA聚合酶,DNA模板约75 ng。PCR扩增反应在Biometra PCR仪上进行。扩增程序:94℃预变性4 min;94℃变性45 s,64℃退火40 s,72℃延伸45 s,每个循环将退火温度降低0.5℃,10个循环;94℃变性45 s,58℃退火40 s,72℃延伸 40 s,22个循环;72℃延伸5 min;PCR产物在4℃条件下保存。PCR产物用PAGE分离,具体方法见王炯等[17]。
1.2.6 数据处理 数据用Excel处理。指标在不同年份间的变异系数计算如下:先用Excel的‘AVERAGE’计算平均值,然后用‘STDEV’函数计算标准偏差,最后计算变异系数。变异系数的计算公式为:变异系数(CV)=(标准偏差/平均值)×100%。相关性分析和主成份分析均用基于R语言的软件(https://www. r-project.org/)完成。相关性用“psych”包的corrtest函数,采用皮尔森方法进行分析;主成分用“FactoMineR”包的PCA函数进行分析。
2 结果
2.1 不同类型砧木资源的种子性状
本试验评价的47份枳均为多核,5年的平均单果种子数为20—44粒。形态观察发现大部分枳种子的种皮光滑,种子饱满(图1)。47份枳中,种子饱满和中等饱满的比例分别为74.5%和25.5%,5年的平均千粒重为141.6—266.2 g。从胚型上看,枳的平均单胚比例为5.2%—99.6%,平均多胚比例为0.4%—94.8%。根据胚型分类标准,47份种质中单胚、多胚和混胚的比例分别为4.3%、2.1%和93.6%(表1,图2-A、2-B)。变异系数是衡量各观测值的变异程度。枳种质多年评价结果显示,种子千粒重的变异系数较小,为3.45%—27.64%,单果种子数的变异系数为2.2%—38.81%,多胚比例的变异系数为3.20%—58.28%,单胚比例的变异系数为0.8%—127.3%(表2、3)。10份枳杂种为较多核和多核,平均单果种子数为8—33粒。枳杂种种子的种皮表面光滑,种子较充实。10份枳杂种中,有6份种子饱满,4份种子中等饱满,其平均千粒重为135.7—239 g,平均单胚比例为0—87.5%,平均多胚比例为12.5%—100%。枳杂种种子的多胚和混胚均为5份,各占50%的比例(图1-A、1-B,表1)。种子千粒重的变异系数为5.66%—27.48%,单果种子数的变异系数为4.72%—76.48%,多胚比例的变异系数为0—12.9%;单胚比例的变异系数为14.14%—84.85%(表2、3)。
12份香橙为较多核和多核,平均单果种子数为12—36粒。香橙种子表面较光滑,种子较充实(图1-E、1-F)。12份香橙中,种子饱满和中等饱满分别所占比例为75%和25%。种子的平均千粒重为147.3—226.3 g,平均单胚比例为0—26.3%,平均多胚比例为73.8%—100%,其中多胚和混胚所占比重分别为58.3%和41.7%(表1,图2-E)。种子千粒重的变异系数为6.71%—52.51%,单果种子数的变异系数为5.14%—54.39%,多胚比例的变异系数为0—26%,单胚比例的变异系数为0—223.61%(表2、3)。27份包含野生、半野生和地方良种的宽皮柑橘种质的种子特性差异较大,有少核、较多核和多核等类型,平均单果种子数为4—62粒。宽皮柑橘的种子外观差异较大,大部分种子比较小(图1-C、1-D)。种子饱满、中等饱满和不饱满的比例分别为11.1%、63.0%和25.9%,平均千粒重为55—391 g。平均单胚比例为0—99.5%,平均多胚比例为0.5%—100%,多胚、混胚和单胚的比例分别为81.45%、11.11%和7.41%(图2-C、2-D,表1)。宽皮柑橘种子千粒重的变异系数为2.67%—36.09%,单果种子数的变异系数为5.41%—74.44%,多胚比例的变异系数为0—200%,单胚比例的变异系数为1.0%—200%(表3)。
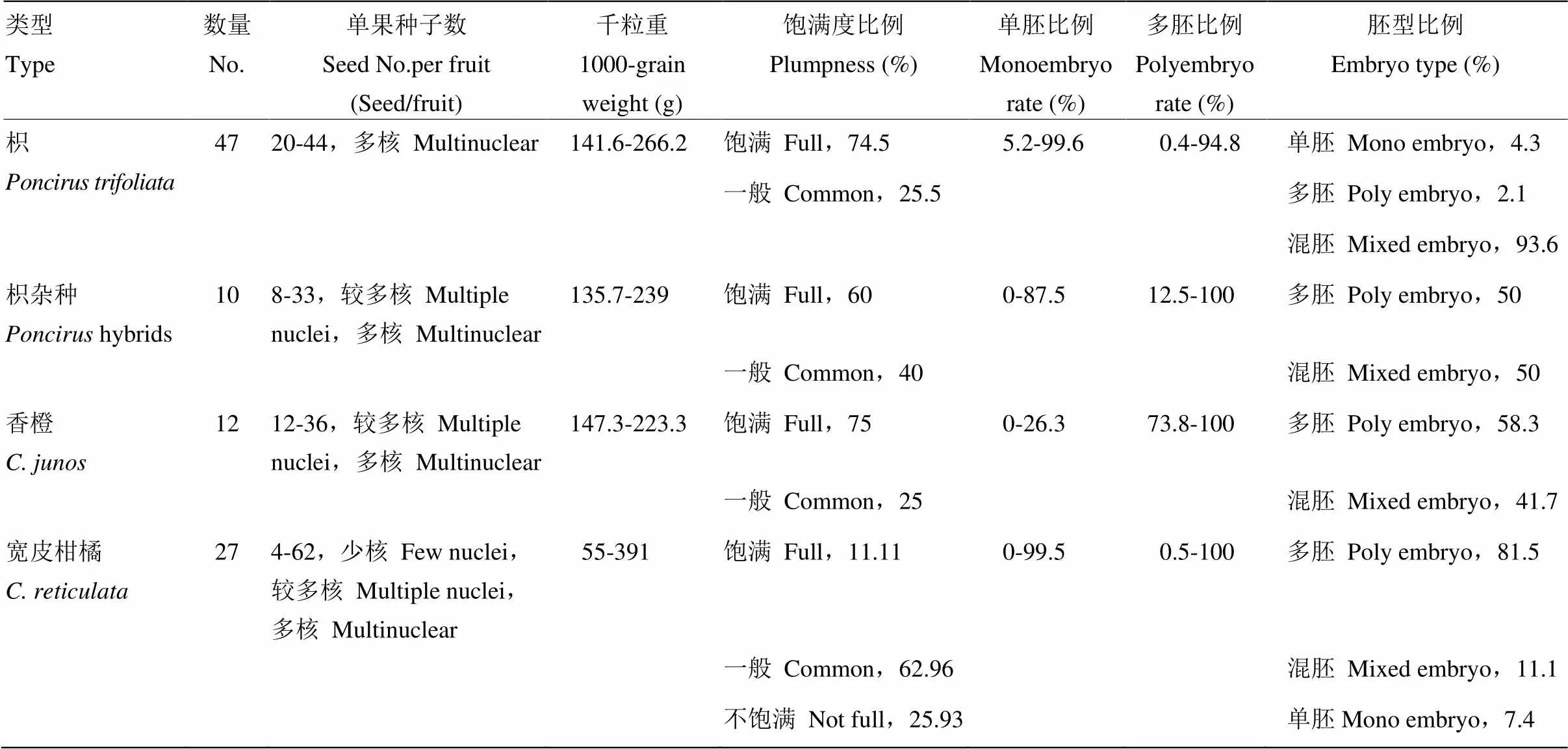
表1 不同砧木种质种子性状指标2013—2017年的5年平均值

A、B:枳杂种,ZZ2-17、ZZ-004;C、D:宽皮柑桔,KPJ-029、KPJ-012;E、F:香橙,XC-004、XC-013
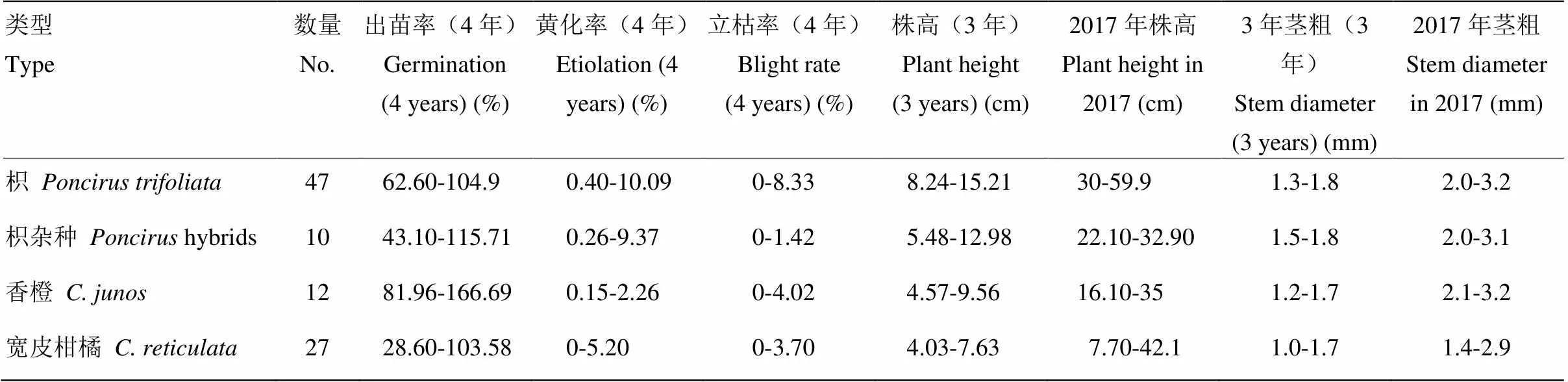
表2 不同砧木种质苗期性状指标多年平均值
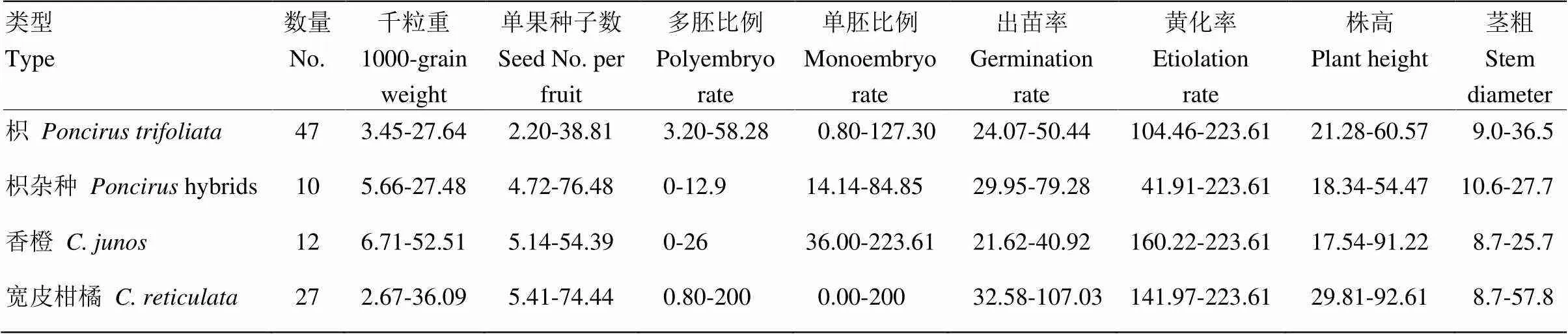
表3 不同砧木种质种子和苗期性状指标的多年变异系数(%)

A、B:枳,Z-027,单胚;Z-043,混胚;C、D:宽皮柑橘,KPJ-011,KPJ-020,多胚;E:香橙,XC-011,混胚;F:香橼,XY-002,单胚
本试验除了评价枳、枳杂种、香橙和宽皮柑橘外,还评价了5份香橼及其杂种、1份酸橙、2份柚及其杂种等其他8份种质。不同类型种质的种子性状和苗期表现差异较大(图3)。柚及柚杂种、香橼的平均单果种子数在47—75粒,为多核;而枸橼和酸橙的平均单果种子数在11—20粒,为较多核。平均千粒重以柚杂种最高,为292 g,而小香橼最低,为68 g。柚杂种的种子饱满,其余几种类型的种子中等饱满,而酸橙种子则不饱满。从胚型上看,柚和2份香橼(图2-F)为单胚,而其余几种为多胚或混胚。

A、B:枳及其杂种Poncirus and its hybrids;A1:ZZ-008;A2:Z-036;A3:Z-024;A4:Z-031;A5:Z-040;A6:Z-042;B1:ZZ-011;B2:Z-030;B3:ZZ-054;B4:Z-028;B5:Z-033;B6:Z-011;B7:Z-014。C、D:宽皮柑橘及其他类型砧木Mandarins and other rootstocks;C1:KPJ-023;C2:KPJ-004;C3:KPJ-029;C4:ZZ-055;C5:KPJ-003;C6:KPJ-005;C7:KPJ-008;C8:KPJ-027;C9:KPJ-013;C10:ZZ-011;C11:KPJ-010;C12:KPJ-018;C13:Y-129;D1:KPJ-028;D2:LM-001;D3:KPJ-012;D4:SC-131;D5:KPJ-002;D6:KPJ-019;D7:KPJ-021;D8:KPJ-009;D9:KPJ-012;D10:XC-006
2.2 不同类型砧木种质的苗期表现
不同枳种质的苗期表现存在差异(图3-A、B)。47份枳4年(因有一年由于特殊原因采果时间提前,导致播种后没有萌发)的平均出苗率为62.6%—104.9%。黄化率为0.4%—10.09%,立枯率为0—8.33%。苗期的生长势方面,苗盘播种4—5个月时的平均株高为8.24—15.21 cm,茎粗为1.3—1.8 mm;栽培槽播种9个月时的平均株高为30—59.9 cm,茎粗为2.0—3.2 mm(表2)。枳出苗率的变异系数为24.07%—50.44%,株高和茎粗的变异系数分别为21.28%—60.57%和9.0%—36.5%,而黄化率的变异系数则较大,为104.46%—223.61%(表3)。枳杂种4年的平均出苗率为69.62%—115.71%,黄化率为0.26%—9.37%,立枯率为0—1.42%。就生长势来看,苗盘播种4—5个月时的平均株高为5.48—12.98 cm,茎粗为1.5—1.8 mm;栽培槽播种9个月时的平均株高为22.1—32.9 cm,茎粗为2.0—3.1 mm(表2)。枳杂种出苗率的变异系数为29.95%—79.28%,株高和茎粗的变异系数分别为18.34%—54.47%和10.6%—27.7%,而黄化率的变异系数则较高,为14.91%—223.61%(表3)。
香橙的苗期表现有明显差异。12份香橙的平均出苗率为81.96%—166.69%,黄化率为0.15%—2.26%,立枯率为0—4.62%。在生长势方面,苗盘播种4—5个月时的平均株高为4.57—9.56 cm,茎粗为1.2—1.7 mm;栽培槽播种9个月时的平均株高为16.1—35 cm,茎粗为2.1—3.2 mm(表2)。香橙种质出苗率的变异系数为21.62%—40.92%,株高和茎粗的变异系数分别为17.54%—91.22%和8.7%—25.7%,而黄化率的变异系数为160.22%—223.61%(表3)。不同宽皮柑橘的苗期表现也有较大差异(图3-C、D)。宽皮柑橘的平均出苗率为28.6%—103.58%,黄化率为0—5.20%,立枯率为0—3.70%。在生长势方面,苗盘播种4—5月时的平均株高为4.03—7.63 cm,茎粗为1.0—1.7 mm;栽培槽播种9个月时平均株高为7.7—42.1 cm,茎粗为1.4—2.9 mm(表2)。宽皮柑橘出苗率的变异系数为32.58%—107.03%,株高和茎粗的变异系数分别为29.81%—92.61%和8.7%—57.8%,而黄化率的变异系数则为141.97%—223.61%(表3)。其他几种种质的平均出苗率为57.89%—113.87%,黄化率为0—6.29%,立枯率为0—1.88%。
2.3 种子和苗期性状指标之间的相关性分析
单果种子数与种子的千粒重、饱满度、单胚比例、幼苗(播种10个月内,下同)株高和茎粗呈极显著正相关,而与多胚比例和胚型呈极显著负相关。千粒重与种子饱满度、单胚比例、幼苗株高和茎粗呈极显著的正相关,而与多胚比例和胚型呈极显著负相关,还与出苗率和黄化率呈显著正相关。种子饱满度与胚型呈极显著负相关,而与单果种子数、千粒重、出苗率、幼苗的株高、茎粗呈极显著正相关。单胚比例与多胚比例、胚型呈极显著负相关,而与幼苗株高呈显著正相关。多胚比例与胚型呈极显著正相关,而与幼苗株高呈显著负相关。胚型与幼苗的株高和5个月幼苗的茎粗呈极显著负相关。幼苗株高与茎粗呈极著正相关,而与胚型呈极显著负相关(表4)。
对种子性状和苗期表现相关指标(黄化率和立枯率除外)进行了主成分分析(图4)。结果表明,这些指标对第一主成分和第二主成分的累计方差贡献率分别为45%和22%,二者合起来的方差贡献率为67%,即包含了原始数据67%的信息,因此可以用这两个主成分代表这104份种质的种子和苗期性状的评价。砧木的单果种子数、种子千粒重、饱满度、胚型、幼苗株高和茎粗对第一主成分的贡献较大,而种子的单胚比例和多胚比例对第二主成分的方差贡献较大。基于种子性状和苗期表现指标,对不同砧木种质进行了主成分分析(图5)。结果表明,单果种子数为多核、种子饱满、幼苗早期生长势旺的枳对第一和第二主成分的方差贡献率均较高,能明显地与其他类型砧木区分,而种子数为少核或较多核、种子不饱满、幼苗生长势较弱的大部分宽皮柑橘种质对第二主成分贡献率较多,而种子数为较多核或多核、种子较饱满、幼苗生长势中等的香橙和枳杂种对两个主成分的贡献率则介于枳和宽皮柑橘之间。
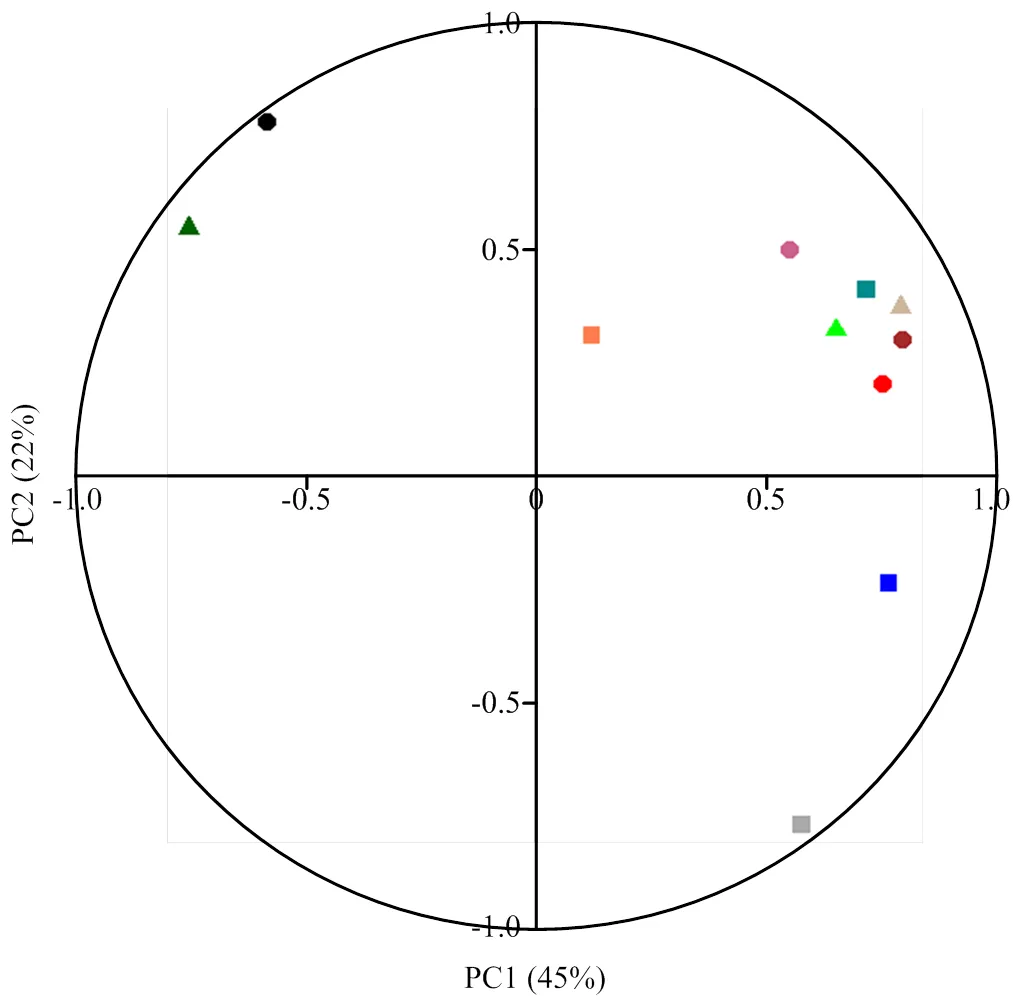
n:单果种子数 Seeds No. per fruits;●:千粒重1000-grain weight;▲:饱满度 Plumpness;n:单胚比例 Monoembryo rate;●:多胚比例 Polyembryo rate;▲:胚型Embryo type;n:萌发率Germination rate;●:2013—2016年株高 Plant height in 2013-2016;▲:2017年株高 Plant height in 2017;n:2013—2016年茎粗 Diameter in 2013-2016;●:2017年茎粗 Diameter in 2017
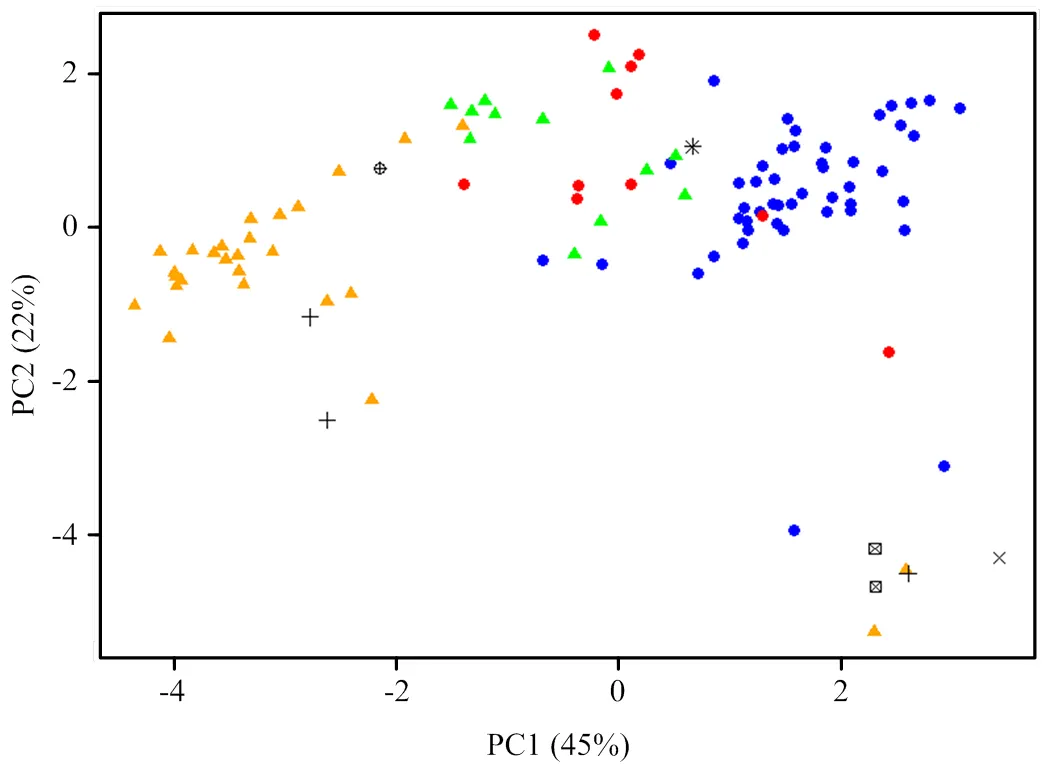
◆:枳 Poncirus;◆:枳杂种 Poncirus hybrid;▲:香橙 C. junos;▲:宽皮柑橘 C. reticulata;+:枸橼杂种 Citron hybrid;×:柚 Citrus maxima;⊕:枸橼 Citron;à:柚杂种 Pummelo hybrid;T:酸橙 Sour orange
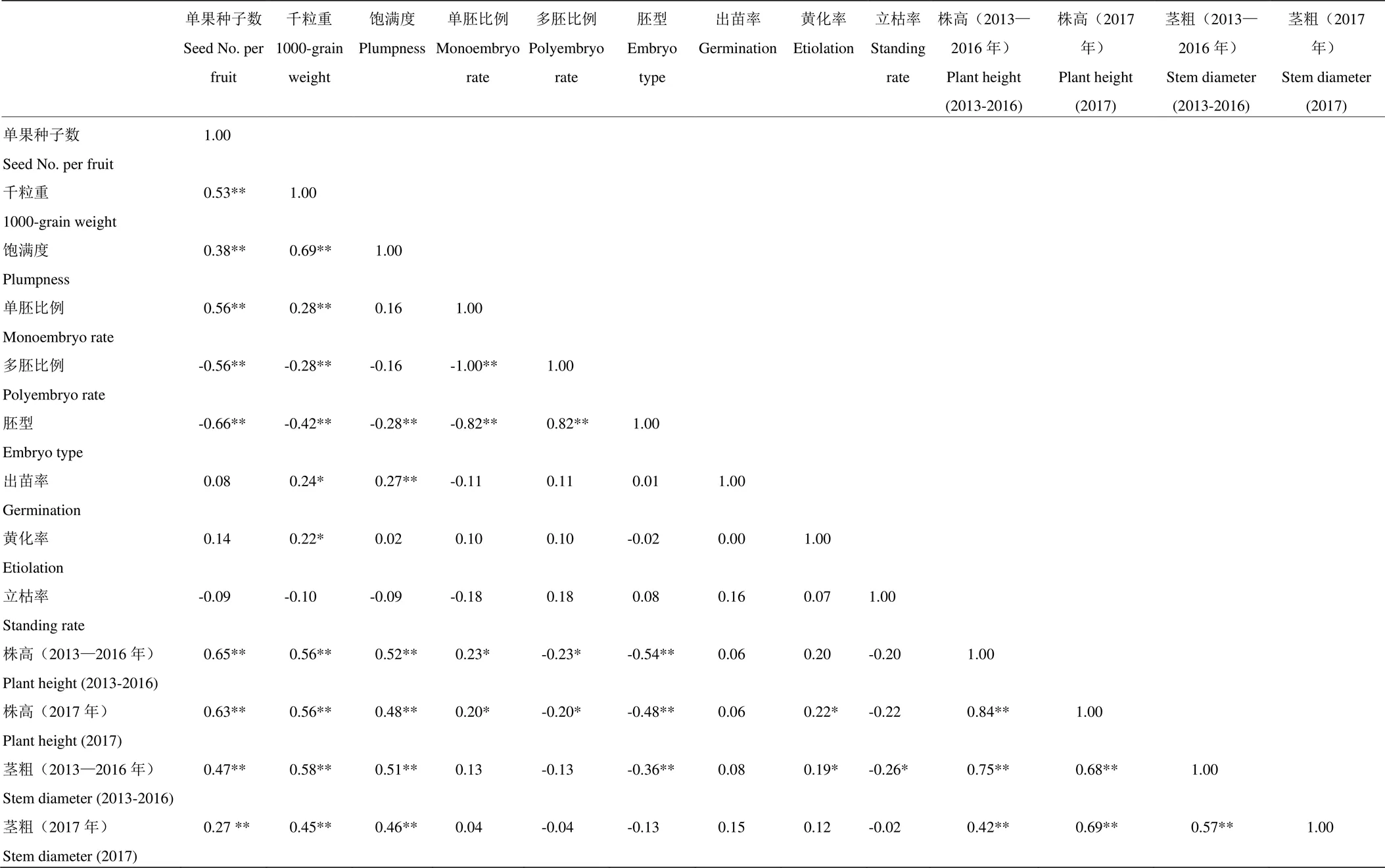
表4 不同砧木种质种子和苗期性状各指标间的相关性分析
*和**分别表示在<0.05和<0.01水平下的相关性
* and ** mean the correlation at<0.05 and<0.01, respectively
2.4 不同胚型种质的MITE插入片段和不同胚型砧木幼苗变异的分子检测
MITE片段插入检测结果显示,5份单胚种质中含RWP结构域的基因启动子中没有检测到MITE片段的插入,在多胚的香橙、宽皮柑橘、枳杂种、柚杂种及酸橙中检测到有MITE片段的插入,这一结果与Wang等[15]、Shimada等[16]的结果一致。但是,在5份无论是单胚还是多胚或混胚的枳种质中均没有检测到MITE片段的插入(图6)。
27对COS标记对8份不同胚型的砧木种质幼苗的一致性分析的结果显示,无论是单胚、多胚还是混胚,其幼苗均有一定程度的变异,因此,在育苗早期根据形态和长势进行除杂是培育优质苗木非常重要的步骤。从胚型上讲,单胚种质KPJ-023幼苗的变异程度最高,而Z-040的变异程度较低,混胚和多胚的变异程度也较低。这些砧木幼苗的变异株率至少为1%(表5)。
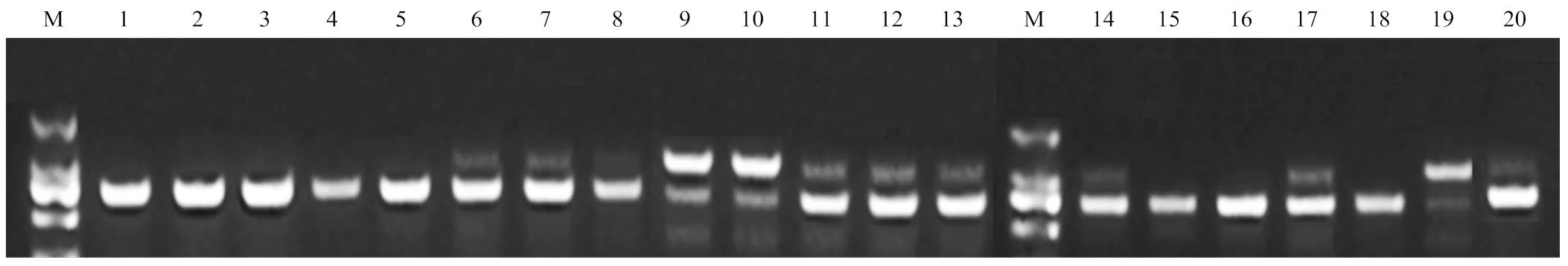
M:标记;1—5:单胚,依次为Y-001(柚);KPJ-012、KPJ-023(尖叶和圆叶莽山野柑);Z-040、Z-027(枳);6—20:多胚,依次为XY-003(香橼);KPJ-001、KPJ-007(宽皮柑橘);XC-013、XC-007、XC-004(香橙);KPJ-011、KPJ-021(宽皮柑橘);Y-129(柚杂种);Z-006、Z-022(枳);ZZ-004(枳杂种);Z-024(枳);ZZ-002(枳杂种);SC(酸橙)
3 讨论
3.1 不同砧木种质的种子性状分析
优良的柑橘砧木不仅要有优良的抗逆性,还要有较强的繁殖能力[12]。与接穗品种需要无核或少核果实不同,砧木品种要求单果种子数量多、质量好、胚型为多胚或混胚以满足大规模培育健壮、一致性好的优质苗木的需要。本次评价的几种主要砧木类型中,以枳、枳杂种和香橙的种子性状较好。总体来看,这3种类型砧木的单果种子数多,为较多核和多核,大部分种子比较饱满。从胚型上看,这3种类型种质中除了有两份枳为单胚外,其余均为多胚或混胚。而所评价的宽皮柑橘种质中,不同种质的种子性状变异较大。单果种子数有少核、较多核和多核,平均千粒重差异也大。在本次测试的104份柑橘砧木资源中,除香橼、柚、KPJ-023、KPJ-012为单胚外,其余的砧木资源均以多胚和混胚为主,这一结果与前人研究结果[5,19]一致。砧木种子的千粒重、单胚比例、多胚比例和单果种子数等多年数据的变异系数均相对较小,一方面说明不同年份之间观测值的离散程度较小,更重要的是说明了这些指标受外在因素的影响较小,遗传稳定性较好。本试验首次在枳种质中发现了两份单胚材料,在应用上,这两份特异资源除可用作砧木育种亲本以提高杂种效率外,也是开展基础研究的好材料。宽皮柑橘砧木种质中,KPJ-023、KPJ-012为单胚,耐寒性强[23],但对盐敏感[24],可以通过杂交育种对其进行遗传改良。
砧木种质的胚型对大规模培育一致性好的优良柑橘苗木非常重要。如果有可靠的分子标记,可以在砧木杂交后代进行早期检测,筛选具有多胚性状的优良单株,这样不仅可以降低育种成本,更重要的是可以缩短选育周期,加快砧木新品种的培育进程。Wang等[15]、Shimada等[16]研究发现,在含有RWP结构域基因的启动子中,MITE片段的插入与柑橘多胚性状紧密相关。为了充分利用MITE片段插入标记进行杂交砧木的早期筛选,对不同胚型砧木种质进行MITE片段插入的检测,大部分种质的结果与Wang等[15]和Shimada等[16]报道的一致,说明可以利用此标记对这些类型砧木杂种的多胚性状进行早期筛选。但是,在检测的5份枳种质中无论是单胚还是多胚均没有检测到MITE插入片段的存在(图6)。究其原因,一方面可能是枳属与柑橘属亲缘关系较远,含有RWP结构域基因的启动子序列差异较大所导致,另外一种可能是枳的多胚性状受其他的机制调控。洪棋斌等[25]根据不同杂交群体的遗传学分析发现,柑橘属和枳属中的无融合生殖(多胚现象)是受位于细胞核内的两对互补的显性基因A1和A2的调控,且其中一对为显性纯合致死效应。因此,关于枳多胚性状的遗传调控值得开展进一步的研究。
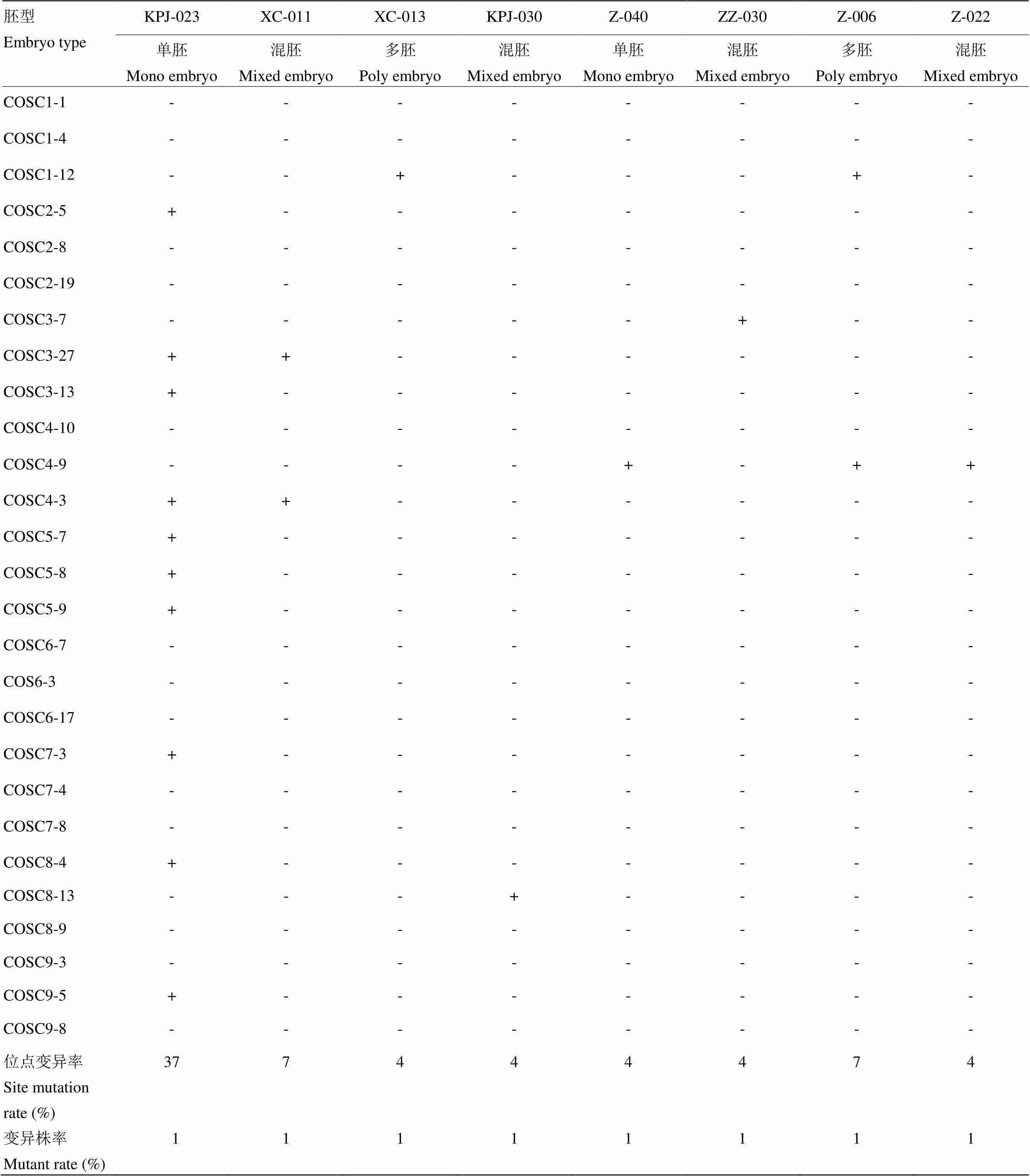
表5 砧木采种树与幼苗一致性检测
表中“-”和“+”分别表示幼苗与种子采种树的遗传背景一致和变异
“-” and “+” mean identity and variation of the seedlings with their seed source tree, respectively
3.2 不同砧木种质的早期苗期性状分析
砧木苗期表现对苗木繁育至关重要。本试验的104份砧木资源中,不同种类砧木以及相同种类不同种质的出苗率差异明显。香橙的出苗率最高,其次是枳和枳杂种,而宽皮柑橘的出苗率则相对较低。本次测试的部分砧木出苗率与Sykes[26]报道的结果较为一致或部分较低,这可能与种质的基因型、种子状态和培养条件有关。
本次评价的104份砧木中,每种砧木类型均出现不同程度和不同比例的黄化现象,其中黄化率<5%的种质居多。关于柑橘黄化的研究,大多数工作都是针对成年树的黄化,主要原因主要有营养元素缺乏、病害、环割过度、土壤酸化板结等,但是对苗期黄化的原因研究较少。有研究认为柑橘苗期黄化可能是受遗传因素和土壤营养状况影响,也有研究认为是种子发育不完全造成的。本试验中砧木幼苗的黄化比例因不同年份而有差异,通过对变异系数分析发现砧木黄化率的变异系数较大,这说明砧木幼苗的黄化可能主要与种子的生理状态、苗床基质的营养状况、管理水平等有关。立枯病是柑橘苗期的主要病害,这4种主要类型的砧木种质中,香橙幼苗对立枯病更为敏感,而枳、枳杂种和宽皮柑橘对立枯病的耐性较好。在生长势方面,对播种10个月内的幼苗进行比较,枳的株高最高,其次是枳和香橙,而宽皮柑橘则最低。
砧木幼苗遗传背景的一致性对培育健康、优质苗木非常重要。在柑橘育苗早期,根据叶片形态和生长势差异去杂是去除遗传背景不一致或天然杂种的有效方法[27]。但要培育优良砧木品种,对幼苗的一致性检测是评价的重要指标。理论上讲,用作砧木的种子应为多胚以保持砧木遗传背景的一致性,因为多胚种子的胚主要由珠心细胞发育而来,其遗传背景与母本相同。本研究对不同胚型砧木资源幼苗遗传背景的分析结果显示,单胚的宽皮柑橘KPJ-023幼苗的变异程度较高,而单胚枳Z-040幼苗的变异程度较低,这说明在开放性授粉条件下,宽皮柑橘KPJ-023与周围柑橘资源的花期相遇,更容易接受外来花粉产生杂种胚,使后代产生较多变异,而单胚枳Z-040幼苗的变异程度比较低,这可能与枳和其他柑橘的花期不遇以及枳所能使用的标记位点变异较少有关。多胚和混胚种质幼苗的变异程度均比较低,其主要原因在于珠心胚多,且珠心胚的位置更靠近营养输导组织,在多胚或混胚种子中,珠心胚的数量和营养优势更加明显,导致杂种胚的退化,因此,播种后的幼苗主要是珠心苗,其与母本在遗传背景上更为一致。但多胚种子的幼苗中也出现了少量的变异株,这可能与体细胞变异或有性多胚有关[28]。无论是多胚还是混胚种子的砧木,种子中均存在较低比例的单胚种子,因此,不能排除非常低频率的杂种胚的可能。
3.3 砧木种子和苗期性状指标的相互关系
砧木的种子和苗期性状指标之间存在紧密的相关性。播种10个月内的幼苗株高和茎粗之间呈显著正相关,而二者与种子的单果种子数、千粒重、饱满度呈极显著的正相关,与胚型呈极显著负相关关系,说明可以通过筛选种子性状优异的砧木选系以提高苗木的质量。但种子胚型和出苗率之间没有相关性,这一结果与Scalon等[29]和Kishore等[30]的研究结果相似。主成分分析结果表明,相关性高的指标对主成分的方差贡献率更高。通过主成分分析,枳、枳杂种、香橙、宽皮柑橘和其他几种类型的砧木种质之间存在明显的差异,这些差异与施文格分类系统中对枳属和柑橘属内种的分类结果比较一致[5]。这一方面说明这些砧木资源在种子和苗期性状指标上的性状差异,另一方面也说明了这些砧木资源在遗传背景、亲缘关系和分类学上的差异,而因为后者的差异才决定了其生物学特性的差异。
4 结论
砧木的种子性状指标和幼苗苗期性状指标高度相关,种子性状优异如单果种子数较多、种子饱满、多胚、千粒重较重的砧木种质,其幼苗的生长势较好。总体来看,枳的种子和幼苗(播种后10个月)的综合性状最好,其次是枳杂种和香橙,宽皮柑橘种子和苗期的综合性状最差。本研究对不同砧木的种子和苗期性状的多年评价结果,可为下一步利用全基因组关联分析(GWAS)中挖掘与柑橘砧木种子数量、胚型及幼苗株高和茎粗相关的候选基因或相关的分子标记研究提供可靠的表型数据。
[1] 周肇基. 中国嫁接技艺的起源与演进. 自然科学史研究, 1994, 13(3): 364-272.
ZHOU Z J. Origin and development of grafting in ancient China., 1994, 13(3): 364-272. (in Chinese)
[2] SNOUSSI H, DUVAL M F, Garcia-Lor A, Belfalah Z, Froelicher Y, Risterucci A M, PERRIER X, JACQUEMOUD- COLLET J P, NAVARRO L, HARRABI M, Ollitrault P. Assessment of the genetic diversity of the Tunisian citrus rootstock germplasm., 2012, 13(1): 16.
[3] ZHU S P, WANG F S, SHEN W X, JIANG D, HONG Q B, ZHAO X C. Genetic diversity ofand phylogenetic relationships with its relatives revealed by SSR and SNP/InDel markers., 2015, 37: 141.
[4] 龚桂芝, 洪棋斌, 江东, 彭祝春, 向素琼. 枳属种质遗传多样性及其与近缘属植物亲缘关系的SSR和cpSSR分析. 园艺学报, 2008, 35(12): 1742-1750
GONG G Z, HONG Q B, PENG Z C, JIANG D, XIANG S Q. Genetic diversity ofand its phylogenetic relationships with relatives as revealed by nuclear and chloroplast SSR., 2008, 35(12):1742-1750. (in Chinese)
[5] 庞晓明,邓秀新,胡春根. 枳属36 份特异种质的AFLP 指纹图谱构建与分析, 园艺学报,2003, 30(4): 394-398.
PANG X M, Deng X X, HU C G. Construction of AFLP fingerprint of 36accessions., 2003, 30(4): 394-398. (in Chinese)
[6] 万良珍, 周开隆. 哈姆林甜橙砧比试验: 1, 枳不同选系的比较.中国柑橘, 1989, 18(4): 3-7.
WAN L Z, ZHOU K L. The comparison test of Hamlin sweet orange on rootstocks: 1. Comparisons of different trifoliate orange selections., 1989, 18(4): 3-7. (in Chinese)
[7] 戴胜根, 李学柱, 马家骐. 枳砧甜橙砧穗组合与黄化的关系研究初报. 中国柑橘, 1986(3): 8-9.
DAI S G, LI X Z, MA J Q. Preliminary research of the relationships between the scion-stock combination of sweet orange onand the etiolation., 1986(3): 8-9. (in Chinese)
[8] 周开隆, 叶荫民. 中国果树志(柑橘卷). 北京: 中国林业出版社, 2009: 145-146.
ZHOU K L, YE Y M.(). Beijing:China Forestry Publishing House, 2009: 145-146. (in Chinese)
[9] Rodríguez-Gamir J, Primo-Millo E, Forner J B, Forner-Giner M A. Citrus rootstock responses to water stress., 2010, 126(2): 95-102.
[10] Zandalinas S I, Rivero R M, Martínez V, Gómez- Cadenas A, Arbona V. Tolerance of citrus plants to the combination of high temperatures and drought is associated to the increase in transpiration modulated by a reduction in abscisic acid levels., 2016, 16: 105
[11] Hussain S, Khalid M F, Saqib M, Ahmad S, Zafar W, Rao M J, Morillon R, Anjum M A. Drought tolerance in citrus rootstocks is associated with better antioxidant defense mechanism., 2018, 40(8): 135.
[12] Wu J X, cAO J Y, su m, feng g z, XU Y H, YI H L. Genome-wide comprehensive analysis of transcriptomes and small RNAs offers insights into the molecular mechanism of alkaline stress tolerance in a citrus rootstock., 2019, 6: 33.
[13] 卿尚模, 易时来, 朱旭荣, 邓烈, 何绍兰, 谢让金. 不同土壤pH值对资阳香橙等4种柑橘砧木苗生长的影响.中国南方果树, 2007, 36(6): 10-12.
QING S M, YI S L, ZHU X R, DENG L, HE S L, XIE R J. The effect of different soil pH on Ziyang xiangcheng and other four kinds of citrus rootstock seedlings., 2007, 36(6): 10-12. (in Chinese)
[14] 解凯东, 王晓培, 谢宗周, 郭文武. 4个柑桔体细胞杂种砧木资源的种子特性评价. 中国南方果树, 2013, 42(5): 53-55
XIE K D, WANG X P, XIE Z Z, GUO W W. Seed traits evaluation of 4 citrus somatic hybrid rootstock germplasm., 2013, 42(5): 53-55. (in Chinese)
[15] Wang X, Xu Y T, Zhang S Q, Cao L, Huang Y, Cheng J F, Wu G Z, Tian S L, Chen C L, Liu Y, Yu H W, Yang X M, Lan H, Wang N, Wang L, Xu J D, Jiang X L, Xie Z Z, Tan M L, Larkin R M, Chen L L, Ma B G, Ruan Y J, Deng X X, Xu Q. Genomic analyses of primitive, wild and cultivated citrus provide insights into asexual reproduction., 2017, 49(5): 765-774.
[16] Shimada T, Endo T, Fujii H, Nakano M, Sugiyama A, Daido G, Ohta S, Yoshioka T, Omura M. MITE insertion- dependent expression ofwith a RWP-RK domain regulates somatic embryogenesis in citrus nucellar tissues.y, 2018, 18: 166.
[17] 王炯, 龚桂芝, 彭祝春, 李一兵, 王艳杰, 洪棋斌. 基于COS Marker 分析柑橘属及其近缘、远缘属植物的遗传与进化. 中国农业科学, 2017, 50(2): 320-331.
Wang J, Gong G Z, Peng Z C, Li Y B, Wang Y J, Hong Q B. Genetic and phylogenetic relationships among citrus and its close and distant relatives based on COS Marker., 2017, 50(2): 320-331. (in Chinese)
[18] Tolly I.. Openbook Howden Print & Design (ISBN 978-0-646- 52065-0), 2017: 148-151.
[19] 中国农业科学院柑桔研究所. 柑桔单胚类型的调查. 柑桔科技通讯, 1975(4): 1-5.
Citrus Research Institute, Chinese Academyof Agricultural Sciences. Survey of mono-embryonic citrus genotypes., 1975(4): 1-5. (in Chinese)
[20] 戴胜根. 培育柑桔优质嫁接苗的关键技术. 中国柑桔, 1995, 24(2): 26-27.
DAI S G. The key techniques of propagating high quality citrus nursery trees., 1995, 24(2): 26-27. (in Chinese)
[21] 王炯. 基于COS Marker构建柑橘连锁图谱及作图群体的光合特性研究[D]. 重庆: 西南大学, 2017.
Wang J. Citrus linkage map construction based on COS marker and photosynthetic characteristics research of the mapping population [D]. Chongqing: Southwest University, 2017. (in Chinese)
[22] 朱世平, 江东, 洪棋斌, 龚桂芝, 刘小丰, 赵晓春. 柑橘砧木育种研究进展. 中国南方果树, 2013, 42(2): 30-34.
Zhu S P, Jiang D, Hong Q B, Gong G Z, Liu X F, Zhao X C. The research progress of citrus rootstock breeding., 2013, 42(2): 30-34. (in Chinese)
[23] Li w B, Zhang y N, Liu g G, He s W. Morphological and isozymic analysis of open-pollinated wild citrus rootstock seedlings//. Guangzhou, 1990: 253-259.
[24] 朱世平, 陈娇, 刘小丰, 曹立, 陆智明, 赵晓春. 15种柑橘砧木出苗期耐盐碱性评价. 西南大学学报(自然科学版), 2014, 36(6): 47-52.
ZHU S P, Chen J, Liu X F, Cao L, Lu Z M, Zhao X C. Evaluation of salinity and alkalinity tolerances of 15 citrus rootstocks by in vitro culture.,2014, 36(6): 47-52. (in Chinese)
[25] 洪棋斌, 向素琼, 陈克玲, 陈力耕. 两对互补的显性基因控制着柑桔属和枳属的无融合生殖. 遗传学报, 2001, 28(11) :1042-1047.
Hong Q B, Xiang S Q, Chen K L, Chen L G. Two complementary dominant genes controlling apomixis in genusand., 2001, 28(11): 1042-1047. (in Chinese)
[26] Sykes S R. Characterization of citrus rootstock germplasm introduced as seeds to Australia from the People’s Republic of China.,2011, 127(3): 298-304.
[27] Xiang C, Roose M L. Frequency and characteristics of nucellar and zygotic seedlings in 12 citrus rootstocks., 1988, 37: 47-59.
[28] 洪棋斌, 李喜庆, 江东, 陈竹生, 向素琼, 龚桂芝, 叶庆亮. 单胚清见橘橙×椪柑产生高比例多胚种子及多胚有性的分子检测. 中国农业科学, 2003, 36(7): 829-833.
HONG Q B, LI X Q, JIANG D, CHEN Z S, XIANG S Q, GONG G Z, YE Q L. High ratio of polyembryonic seeds in monoembryony Kiyomi × Ponkan and molecular evidences against asexual origins for polyembryonies., 2003, 36(7): 829-833. (in Chinese)
[29] Scalon S D P Q, Mussury R M, Rigoni M R, Filho H S. Initial growth of(Pasq.) A. Robyns seedlings under shading conditions., 2003, 27: 753-758.
[30] Kishore K, Monika N, Rinchen D, Lepcha B, Pandey B. Polyembryony and seedling emergence traits in apomictic citrus., 2012,138: 101-107.
Seed Traits and Seedling Performances of Different Types of Citrus Rootstock
ZHU ShiPing1, WANG FuSheng1, CHEN Jiao2,3, YU Xin1, YU Hong1, LUO GuoTao1, HU Zhou1, FENG JinYing1, ZHAO XiaoChun1, HONG QiBin1
(1Citrus Research Institute, Southwest University/Chinese Academy of Agricultural Sciences/ National Citrus Engineering Research Center, Chongqing 400712;2College of Horticulture and Landscape Architecture, Southwest University, Chongqing 400716;3Bureau of Agriculture and Rural Affairs of Fushun County, Fushun 643200, Sichuan)
【Objective】Good rootstock could promote plant growth, improve fruit yield and quality, and enhance stress tolerance and adaptability. Currently in China, mixed genotype of seeds is generally used for propagation of nursery trees. Variable germination, growth, and stress tolerance of seedlings are the common problems significantly affecting the quality of nursery trees, also the performance of orchard plants. This research was aimed to study the key parameters related to the quality of seedlings in different genotypes of rootstocks, to establish the standards for evaluation of rootstock and provide guidelines for selection of superior rootstock. 【Method】 104 different citrus rootstock germplasm accessions were evaluated in five consecutive years on their seed characteristics, such as seed number per fruit, plumpness of seeds, 1 000-grain weight, embryonic type, and the seedling performances, such as germination rate, occurrence of etiolation and blight, and the growth vigor. The uniformity of the seedlings of some rootstock accessions were identified by COS (Conserved Ortholog Sequences) markers. Association of MITE (Miniature inverted-repeat transposable element) insertion with embryonic type was also investigated in some rootstock accessions. 【Result】 (1) Fruits ofand its hybrids, as well asaccessions were highly multi-seeded and multi-seeded. Most of the seeds were plumpy, polyembryonic and mixed-embryonic type. The 1000-grain weight of most accessions was more than 200 g. The seed numbers per fruit in mandarins were highly variable in the range of low-seeded, multi-seeded and highly multi-seeded. Most seeds were medium plumpy and polyembryonic. The 1 000-grain weight in approximately half of the mandarin accessions was less than 100 g. (2) Seedling performances differed among different types of rootstocks.demonstrated the highest frequency of germination, and lower etiolation rate, but the seedlings were sensitive to blight disease. The germination rates ofand its hybrids were lower than that ofbut the etiolation rate was higher. Their seedlings were tolerant to blight disease. The germination rates of most mandarin accessions were comparatively low, and the rates of both etiolation and blight were also low. Comparisons of seedling growth at 10 months after sowing,showed the strongest growth vigor, followed by its hybrids and, while those of mandarins were the least measured by both seedling height and stem diameter. (3) Correlation analysis revealed that except for the occurrences of etiolation and blight, parameters between the traits of seeds and seedlings such as seeds per fruit, 1000-grain weight, plumpness, the mono-embryo rate, poly-embryo rate, embryonic type, the height and the stem diameter of seedlings were tightly correlated at the levels of high significance (<0.01) or significance (<0.05). The contribution of each individual parameter to the overall variation was highly different. PCA (principle component analysis) could clearly distinguish the genetic differences between the different types of rootstocks. The coefficient variations of the parameters except for etiolation rate were not significantly variable among different years, indicating those traits were comparatively stable. (4) Results of COS marker showed that mono-embryonic seedlings of mandarin were less uniform than that of mono-embyonic seedlings ofand all the poly-embryonic and mixed-embyonic accessions. MITE insertion was detected in polyembryonic accessions of mandarin,andhybrids, but did not in monoembryonic accessions of mandarin and pummelo. However, MITE insertion did not observed either in monoembryonic or polyembryonic accessions of some. 【Conclusion】Parameters between seed traits and seedling performances were highly correlated. These results indicated that the high quality of seedling was greatly associated with good characters of seed. Measured by overall performance, the characters of both seeds and seedlings ofwere the best, followed byhybrids andthose of mandarins were the worst.
citrus rootstock; seed trait; embryonic type; seedling performance; uniformity
2019-07-09;
2019-09-19
国家重点研发计划(2018YFD0201503)、现代农业产业技术体系项目(CARS-26)、中央高校基本业务费(XDJK2016B024)、国家柑桔工程技术研究中心开放课题资助项目、重庆市现代山地特色高效农业技术体系创新团队建设计划(晚熟柑桔脆李技术体系)、西南大学双一流学科建设项目、西南大学博士/引进人才计划基金(SWU117038)
朱世平,E-mail:zhushiping@cric.cn。通信作者洪棋斌,E-mail:hongqibin@cric.cn
(责任编辑 赵伶俐)
