新疆开都河长身高原鳅的年龄与生长的关系
2020-02-14金珊珊王新月陈生熬刘茂春谢从新
金珊珊 , 王新月, 林 欣,陈生熬, 刘茂春,谢从新
(1. 塔里木大学动物科学学院,新疆阿拉尔 843300;2. 华中农业大学水产学院,武汉 430070;3. 四川律贝生物科技有限公司,成都 610011)
0 引 言
【研究意义】开都河发源于天山南麓中部的伊连哈比尔尕山,经由孔雀河流至博斯腾湖,最终流入塔里木河流域,全长约500 km,流域面积2.2×104km2,流经巴音郭楞蒙古自治州和静、焉耆、博湖三县,塔里木河水系主要支流之一;曾经是新疆裸重唇鱼(Gymnodiptychusdybowskii)和斑重唇鱼(Diptychusmaculates)、长身高原鳅(Triplophysatenuis)等主要栖息地之一;由于外来物种带入,水域生态劣化,土著鱼类渔业资源锐减,长身高原鳅等特有土著鱼类濒危[1-4]。【前人研究进展】开都河相关研究最早见于20世纪60年代测绘工程等,80~90年代水利建设等研究[5-9];但对于渔业的研究,主要在2000年以后该水系裂腹鱼类和高原鳅等土著鱼类区系、水生生物等渔业资源调查、以及外来鱼类组成等生态学研究,以及水质理化分析等多方面报道[4, 9, 10]。【本研究切入点】近年来,对于该流域中土著鱼类的报道较少[11-13]。且未见长身高原鳅等土著鱼类系统生物学研究报道。研究开都河长身高原鳅年龄与生长的关系。【拟解决的关键问题】2017~2019年对开都河长身高原鳅的年龄与生长关系进行分析,研究南疆高旱地区土著鱼类的生长特性及特有生态系统,为渔业可持续发展打基础。
1 材料与方法
1.1 材 料
2017~2019年,在开都河上游河段,N42。32,~E85。31,分4季,用刺网和围网(2 a=2.0 cm)采集长身高原鳅139尾(雌性83尾,雄性56尾),现场生物学测定并解剖,取出耳石、3~8节脊椎骨和鳃盖骨,保存带回实验室处理。
1.2 方 法
参照殷名称[14],对长身高原鳅体长(L)、体重(W)、头长(LH)、体高(WH)、体宽(BW)、肠长(LI)、性腺重(WG)等指标进行测量(长度精确至 0.01 cm,质量精确至 0.01 g)。
依据殷名称方法[14],采集耳石和第3~8节脊椎骨,用清洗、脱脂(KOH浸泡5~10 min)、清洗、透亮(二甲苯透明)、观察(2人以上)、鉴定年龄。
1.3 数据处理
依据殷名称等[14],通过幂函数W=aLh对体长和体重拟合分析;
采用公式K=100(W/L3)求得丰满度,观测其生长性状;
采用GSI=100(W/WG)求得成熟系数,观察其性成熟分期;
生长方程,采用von Bertalanffy(1933)生长方程描述年龄与生长特性:
Lt=Lt(1-e-k(t-t0)),Wt=Wt(1-e-k (t-t0));
通过 Ford方程和 Beverton法求得L∞、k和t0;
采用SPSS17.0和ORIGIN9.0,并用均值和标准差(Mean±SD)表示。
2 结果与分析
2.1 年龄结构和体长/体重分布
研究表明,长身高原鳅年龄范围为1~6,均值为3.06±0.11+,不符合正态分布(SPSS17.0检验,Sk=0.622,Ku=-0.504);其中2+为优势年龄个体,占36.00%;3+,占28.10%,4+和5+仅占11.50%和14.40%。
长身高原鳅体长范围为4.78~13.03 cm,均值为(8.90±0.13 )cm,不符合正态分布(SPSS17.0检验,Sk=0.180,Ku=-0.055);其中优势体长10.03 cm,占2.90%。
长身高原鳅体重范围为1.83~18.74 g,均值为(6.65±0.31 )g,不符合正态分布(SPSS17.0检验,Sk=0.995,Ku=0.187),其中优势体重3.40和3.76 g,仅占2.90%多数个体所占比例基本一致,仅为0.7%。
长身高原鳅肠长范围为3.45~16.83 cm,均值为(9.28±0.16 ) cm,不符合正态分布(SPSS17.0检验,Sk=0.662,Ku=3.07),其中优势肠长9.52 cm,仅占2.90%,多数个体所占比例基本一致,仅为0.7%。
采集样本中,长身高原鳅最小1龄,最大6龄,各自体长和体重分别是6.80 cm和2.38 g,以及11.25 cm和体重15.19 g。图1
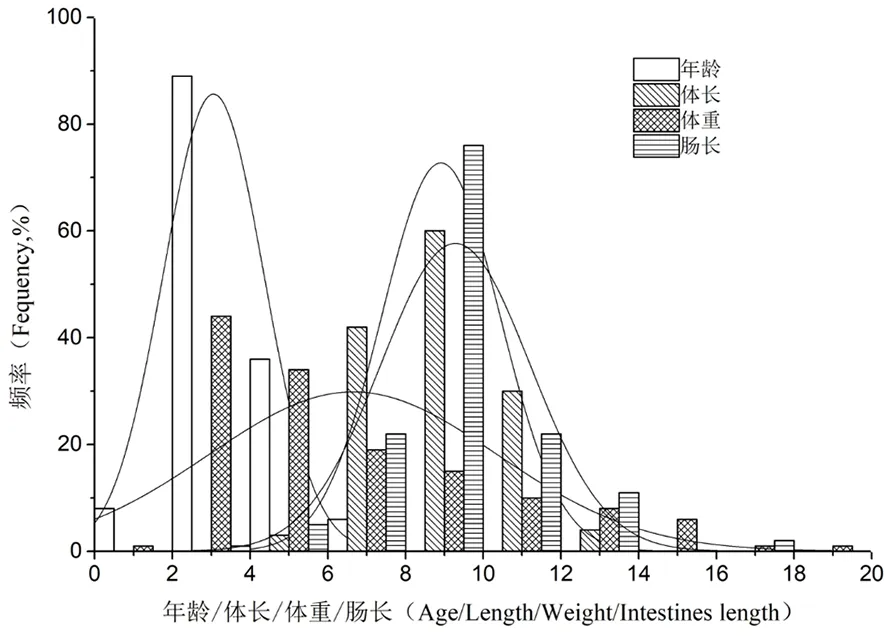
图1 长身高原鳅年龄/体长/体重/肠长分布
Fig. 1 Distributions of the standard age/length/weight/intestine length frequency ofTriplophysatenuis
长身高原鳅雌性个体年龄结构可知,其个体的年龄范围为2~5,均值为2.63±0.09+,不符合正态分布(SPSS16.0检验,Sk=1.232,Ku=1.14);其中2+为优势年龄个体,占51.80%;3+,占34.10%,4+和5+仅占8.2%和3.5%。
长身高原鳅雄性个体年龄结构可知,其个体的年龄范围为1~6,均值为3.70±0.21+,不符合正态分布(SPSS16.0检验,Sk=-0.356,Ku=-1.058);其中5+为优势年龄个体,占20.00%;3+,占11.80%,2+和6+均占7.10%。图2
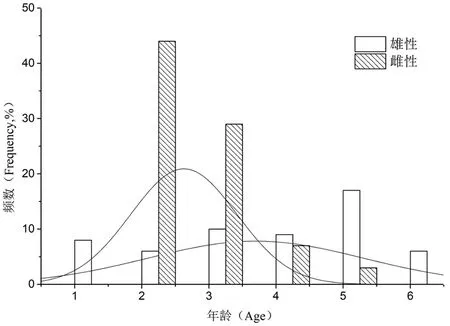
图2 长身高原鳅雌雄个体年龄分布
Fig. 2 Distributions of the standard age frequency ofTriplophysatenuis
2.2 生长分析
2.2.1 体长与体重的关系式
对长身高原鳅(n=139)体长与体重回归分析得:
W总=0.013 3L2.809 4(R2=0.745 9)。
研究表明,开都河中,长身高原鳅多数个体体长分布范围在7.00~10.00 cm。W=0.013 3L2.809 4(R2=0.745 9)关系式中b=2.809 4<3(P<0.05),属于异速生长。图3
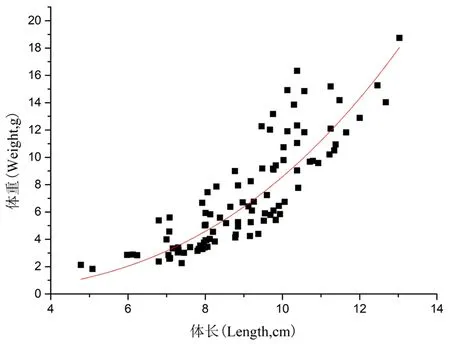
图3 长身高原鳅体长与体重关系
Fig. 3 Length-weight relationships ofTriplophysatenuis
研究表明,长身高原鳅雌性个体多数体长分布范围在7.00~10.00 cm。W=0.005L2.720 6(R2=0.807 7)关系式中b=2.720 6<3(P<0.05),属于异速生长。长身高原鳅雄性个体多数体长分布范围在7.00~10.00 cm。W=0.019L2.371 3(R2=0.763 0)关系式中b=2.371 3<3(P<0.05),属于异速生长。图4
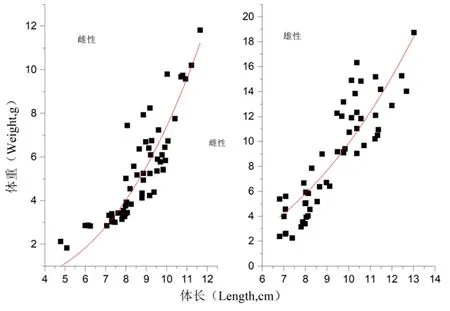
图4 长身高原鳅(雌雄)体长与体重关系
Fig. 4 Length-weight relationships ofTriplophysatenuis(♂♀)
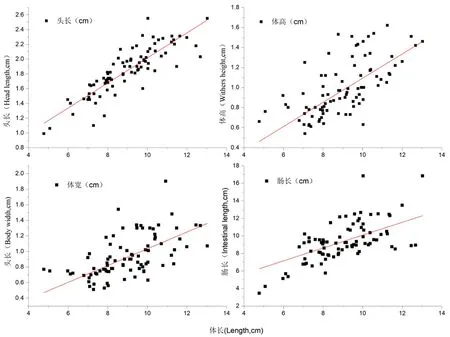
图5 长身高原鳅体长与头长/体高/体宽/肠长的关系式
Fig.5 Length- head length/ Withers height/Body width/intestine length relationships ofTriplophysatenuis
2.2.2 体长与其他指标的关系
拟合长身高原鳅(n=139)体长和头长、体长和体高、体长和体宽、体长和肠长等关系进行回归方程。
LH=0.169L+0.320(R2=0.846 3);体长与头长相关性较高,呈现直线相关。
WH=0.120L-0.110(R2=0.688 1);体长与体高相关性较低,呈现直线相关。
BW=0.107L-0.040(R2=0.605 9);体长与体宽相关性较低,呈现直线相关。
LI=0.730L-2.74(R2=0.981 8);体长较为稳定,范围在45.00~50.00 cm。与肠长相关性较高,二者直线增长趋势明显;其中肠道和体长之比范围浮动在(1.05±0.18),其中小于3,大于1,符合杂食性鱼类摄食习性。图5
2.2.3 生长方程
通过长身高原鳅(n=139)实测体长,退算求得渐进体长L∞=23.75 cm,k=0.64,W∞=97.42 g,t0= - 0.70,ti=9.14。
求得生长方程:
Lt= 23.75 (1-e-0.64 (t + 0.7)) ;
Wt= 97.42 (1-e-0.64 (t + 0.7))2.809 4。
对生长方程进行一阶求导,得到生长速度方程:
dL/dt=15.20e-0.64 (t + 0.7);
dW/dt=175.16e-0.64 (t + 0.7)
(1-e-0.64 (t +0.7)1.809 4。
对生长方程进行二阶求导,得到生长加速度方程:
dL2/dt2=-9.73e-0.64 (t + 0.7);
dW2/dt2= 112.10e-0.64 (t + 0.7)(1-e-0.64 (t + 0.7)0.809 4(2.809 4×e-0.64 (t + 0.7)-1)。
长身高原鳅的体长与体重渐近生长曲线相似。体长与体重的生长速度呈下降性正相关,但体长生长速度趋势缓于体重生长速度,生长加速度也表现出与生长速度类似的趋势,但体重生长加速度在0~2龄之间表现出直线下降趋势,2龄处再行上升与体长加速度一致。体长生长曲线随年龄持续增大。长身高原鳅拐点年龄ti为0.914,所对应长身高原鳅体长和体重分别为:15.29 cm和28.30 g。体重生长速度增长趋势平稳,当到达拐点年龄后,平稳增长。图6
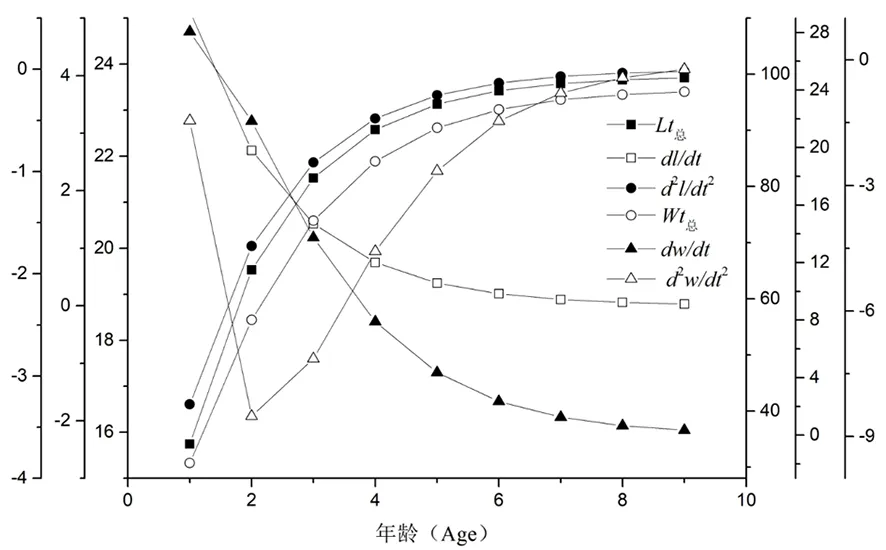
图6 长身高原鳅体长和体重von Bertalanffy生长曲线/生长速度/生长加速度
Fig. 6 Von Bertalanffy growth/growth rate/growth acceleration of the standard length and weight ofTriplophysatenuis
2.2.4 长身高原鳅成熟系数和丰满度
研究表明,GSI=27.20±1.80。从长身高原鳅的性腺成熟度中可以看出,其繁殖盛期主要在5~6月,持续时间从4~8月。图7
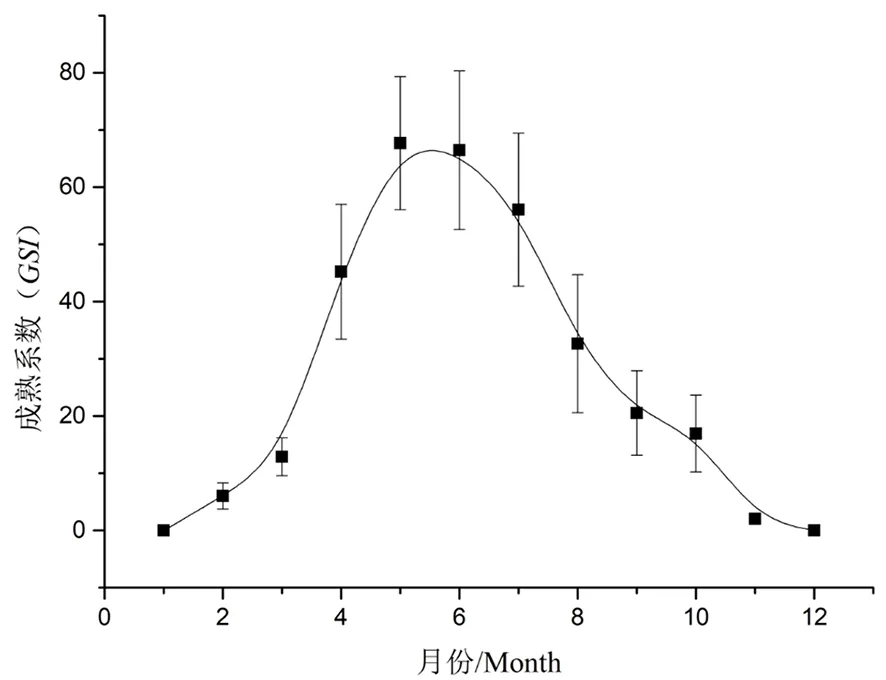
图7 长身高原鳅的成熟系数
Fig. 7 GSI ofTriplophysatenuisfrom Shangyou reservoir
研究表明,K=0.88±0.26。丰满度出现正态分布趋势,每年中出现1次峰值,长身高原鳅产卵类型为1次产卵,和其他水系高原鳅是一样的。 图8
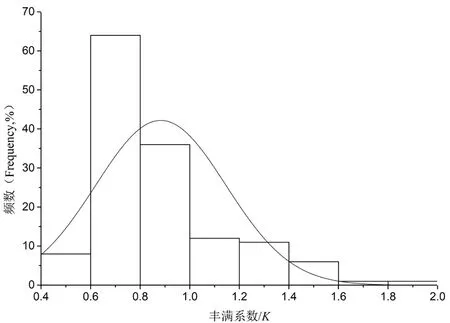
图8 长身高原鳅的丰满度
Fig. 8 Fullness ofTriplophysatenuis
3 讨 论
新疆塔里木河流域,由9大水系、144条河流组成,水量较大,属于塔里木河流域中上游主要支流之一,发源于天山南麓中部的伊连哈比尔尕山,流经巴音郭楞蒙古自治州流至孔雀河入博斯腾湖[15, 16];夏季最高水温为16℃,最低2℃,EC=282.96 Us/cm,BOD= 15 mg/L,DO=69.57%,pH=7.78,SAL=9.36 mg/L,水温较低,水量充沛,是高原裂腹鱼类和高原鳅属鱼类栖息的好环境[17-20];开都河独特的河流生态环境,维系着新疆面积最大的天鹅湖湿地及草原[1, 2, 16]。塔里木河流域渔业产值主要在塔里木河上游段平原型水库渔业[18, 21-24];开都河渔业的主要在常规鱼类调查捕捞及鱼类区系的划分,以及博斯腾湖渔业等[4, 9]。
开都河流域中,由于该区域主要以棉花等为主,耕地面积增加,灌溉水量逐渐增大,压盐碱水不同程度进入到干流及孔雀河等,使得盐碱升高,饵料生物缺乏,这与塔里木河流域诸多水库水源来自天山融雪,高盐碱灌溉水进入塔里木河等密切相关,进而影响鱼类生长[25, 26]。开都河鱼类区系及分布通过稳定和发展,以及人为引种等因素影响下,有较明显区域特征,再者由于河流水温低以及水利枢纽等,使得出山口后河段的鲤和鲫等外来鱼类,和出山口以上河段的裂腹鱼和高原鳅等土著鱼类出现了空间分布的明显界限[4]。
研究发现,长身高原鳅栖息环境主要在塔里木河流域,例如塔里木河上游段、阿克苏河以及叶尔羌河流域等水域;其中长身高原鳅体长与体重关系式中,塔里木河上游段个体[27]W=0.098L2.094、阿克苏河[28]个体W=0.009L2.966 6,其中和研究中开都河个体W总=0.013 3L2.809 4相互比较,b值分别是2.094、2.966 6和2.898 4,a值分别是0.098、0.009和0.013 3;其中阿克苏河长身高原鳅生长速度最接近匀速生长,塔里木河上游段个体明显异速生长,塔里木河上游段鱼类肥满度高,异速生长中,长身高原鳅能量可能主要集中在繁衍生息[14]。有研究发现,长身高原鳅的体长与肠长关系式为LI=0.730L-2.74(R2=0.981 8),其中肠/体比,1<1.05<3,符合杂食性鱼类摄食习性[17, 27, 28]。
依据Von Bertalanffy(1938)拟合的生长方程比较[14]:长身高原鳅研究中L∞=23.75 cm,k=0.133,W∞=97.42 g,t0= - 0.70,ti=9.14;与东方高原鳅相比较,L∞=15.16 cm,W∞=31.49 g,t0= - 0.017,ti=8.19,其中ti变化趋势明显,其他指标类似,这与不同水系有关[29];麻尔柯河高原鳅L∞=17.31 cm,k=0.159 7,W∞=60.75 g,t0= - 0.532 8,ti=6.25;不同水系长身高原鳅的生长与不同水域的水源及水温均较低有关[30]。开都河长身高原鳅的拐点年龄大于其余水系拐点年龄,表现出生长速度慢,机体偏瘦,这与近年来人们对土著野生鱼类的奢求,以及病态捕捞等,有关使得盲目的引入鱼类品种,造成生态位竞争,未曾达到鱼类的最适生长理论[14]。
长身高原鳅生长发育中,阿克苏河繁殖时间主要在4~5月,塔里木河上游段繁殖时间在3~7月,研究中4~8月,这其中包含了5月生产盛期,三者水域相一致,但由于水域水温、浮游生物及泥沙等不同,持续月份有所差异[17, 27, 28]。长身高原鳅的雌雄鉴别方法和其他高原鳅属鱼类一致[31];在人工繁殖中,叶尔羌河流域个体在自然水体中长身高原鳅雌雄比的1∶1.5,阿克苏河中则为1∶1.1,这与塔里木河上游段个体的雌雄比为1∶1.63相似,相比较研究中为1.4∶1则差异显著(P<0.05)[17, 27, 28]。长身高原鳅繁殖个体大小中,叶尔羌河流域为体长7.1~10.2 cm,体重6.3~9.8 g;阿克苏河为5.4 ~6.6 cm,体重1.4~2.7 g,与研究中相比(5.08~11.65 cm,体重1.83~11.82 g)体长与体重均值大于叶尔羌河流域和阿克苏河[17, 27, 28]。长身高原鳅的成熟系数比较中,阿克苏河[28]为21.4,研究中为27.20±1.80,两者差异显著(P<0.05),这与栖息水域环境密切相关。
4 结 论
塔里木河流域开都河,自中天山融雪汇入孔雀河流至博斯腾湖,造成水域环境不同,水质理化指标不同,浮游生物不同,长身高原鳅年龄与生长差异显著(P<0.05)。开都河各个河段水温低,饵料丰度低,栖息于开都河的长身高原鳅生长慢,年龄组成1~6龄,范围较大,年龄均值为3.06±0.11+,优势年龄仅为2+,结构单一,不符合正态分布趋势(SPSS16.0检验,Sk=0.622,Ku=-0.504)。从体长与体重关系式表现出异速生长,尾柄较其他高原鳅长,与其肠长相关性较大,丰满度表现为较低值0.88±0.26,生长周期较长,拐点年龄高达9+,成熟系数GSI=27.20±1.80,存在小型化趋势等显著特征。
