河北省葡萄座腔菌科菌物系统分类研究
2020-02-12王庆灵于贵朋邢军超崔建州赵嘉平
王庆灵,于贵朋,邢军超,崔建州,赵嘉平
(1.中国林业科学研究院林业新技术研究所,北京 100091;2.中国林业科学研究院热带林业试验中心,广西 凭祥 532600;3.河北省塞罕坝机械林场,河北 承德 068450;4.河北农业大学 林学院,河北 保定 071001)
树木溃疡病是引起多种树木流水流胶、组织坏死和在病斑周围形成愈伤组织等症状的病害总称。葡萄座腔菌科Botryosphaeriaceae 真菌是引起树木溃疡病的主要菌物[1-2]。狭义上,树木溃疡病原真菌是指葡萄座腔菌科真菌,具有寄主植物多样、危害症状复杂、分布广泛等特征[1,3]。树木溃疡病病原真菌危害多种生态及经济林木的发展,特别是对杨树人工林[4-5],已造成严重的生态和经济损失,是我国森林生物灾害之一[6]。在河北省,葡萄座腔菌科真菌曾引发核桃Juglans regia、板栗Castanea mollissima等多种植物的枝干溃疡病,严重时导致整株死亡[7-9]。虽然葡萄座腔菌科真菌的有性型名称更适合形态鉴定,但由于在自然界和培养条件下很少见到有性型,因此鉴定相应真菌的主要依据是较为常见的无性型;然而,某些种类真菌的无性型形态的某些特征会出现交叉重叠现象,且某类的孢子可能随着孢子发育的时间发生变化[10-14]。因此,形态鉴定结合分子鉴定成为葡萄座腔菌科真菌的现代分类方法,有时会考虑病原的致病力和病害症状等[15-21]。笔者从河北省各地的树木上采集到疑似溃疡病的枝干样本,并分离出菌株。然后综合应用培养学、形态学以及rDNA-ITS 系统发育特征对河北省树木溃疡病害的病原菌物进行分类鉴定,其研究结果将为河北省树木枝干溃疡类病害的综合控制奠定坚实的实验基础和理论依据。
1 材料与方法
1.1 试验材料
2011年1—5月份从河北省石家庄、保定、沧州、邢台、邯郸、衡水、承德、张家口等8 个县市广泛采集不同树种、不同病害症状的树木溃疡病枝干样本。将新鲜的有典型发病症状的枝干标本,在室内利用常规组织分离法和菌落边缘菌丝纯化法得到供试菌株[20]。分离和纯化实验均在2% PDA 培养基上进行,共得到73 份试验菌株。
1.2 病原菌的形态鉴定
1.2.1 培养学、形态学特征的获得
本研究试验菌株活化培养采用2%PDA 培养基(马铃薯200 g,葡萄糖20 g,琼脂15 g,水 1 000 mL),25 ℃、黑暗培养4 ~7 d,观察菌落典型特征。采用松针培养基诱导子实体形成,具体方法为:健康、新鲜的西黄松Pinus ponderosa Dougl.ex Laws松针剪成4 ~6 cm 小段,灭菌后放置在凝固的无菌水琼脂培养基上,接入试验菌株PDA 培养物,封上封口膜;28 ℃黑暗培养10 d 左右,去掉封口膜,于室温条件下继续黑光灯光照培养20 d 以诱导子实体的形成,镜检试验菌株的形态学特征。
1.2.2 培养学、形态学特征的观察
取典型的菌株子实体分别做徒手切片,在显微镜下观察病菌的分生孢子和分生孢子器形态特征,并测量其大小。对理想的真菌切片进行拍照并描述。根据试验菌株的培养学性状和形态学特征,采用葡萄牙微生物里斯本大学微生物资源中心(Centro de Recursos Microbiológicos, CREM)的葡萄座腔菌分类检索表(http://www.crem.fct.unl.pt/botryosphaeria_site/key.htm)及相关文献对参试菌株进行初步鉴定[4]。
1.3 真菌的分子生物学鉴定
1.3.1 病原菌DNA 的提取
试验菌株在PD液体培养基(马铃薯200 g,葡萄糖20 g,水1 000 mL)中静置培养7 d,挑出菌丝并用滤纸吸干水分,直接提取DNA 或者-20 ℃ 保存备用。DNA 提取在冰上进行,用冷冻研磨棒充分研磨病原菌物菌丝直至糊状,DNA 提取采用北京博迈德科技发展有限公司的CTAB 植物基因组DNA 提取试剂盒。分光光度计测定DNA 纯度和浓度,4 ℃条件下保存。
1.3.2 病原菌rDNA-ITS 片段扩增及系统发育分析
利用引物对ITS1(5’TCCGTAGGTGAACCTG CGG3’)/LR5(5’TCCTGAGGGAAACTTCG 3’), 聚合酶链式反应(PCR 反应)扩增供试菌株rDNA-ITS序列片段。PCR 反应采用40 μL 扩增反应体系。每个反应体系DNA 模板加入量为 2 μL,上下游引物(浓度为10 μmol/L)各2 μL,2×Taq PCR Master Mix 体系加入量为20 μL,最后用14 μL 灭菌的去离子水补足40 μL。PCR 扩增共40 个循环,退火温度58 ℃,退火时间30 s,扩增循环完成后72 ℃延伸7 min。1.5%琼脂糖凝胶电泳检测PCR 产物。PCR 扩增产物测序分析委托北京博迈德科技发展有限公司,测序引物为ITS1引物。
在NCBI 中使用Blast 在线比对序列,从GenBank 中提取37 个参比菌株rDNA-ITS序列。使用MEGA 5.05 软件中的Muscle,对试验菌株的rDNA-ITS序列和参比菌株rDNA-ITS序列进行同源比对分析;分别采用P-距离、Kimura-2 距离,以邻位连接方法(Neighbour-joining method,NJ)构建系统发育树。
2 结果与分析
2.1 病原菌的培养学、形态学特征及鉴定
本研究从河北省石家庄、保定、沧州、邢台、邯郸、衡水、承德、张家口等8 个市(区、县)25 个不同地点进行树木溃疡病害标本的采集,共分离获得河北省树木溃疡病病原菌物73 株,菌株鉴定结果、寄主植物、采集地点等信息见表1。溃疡病害的寄主植物包括杨树、柳树、苹果、核桃、大叶黄杨、构树、火炬树、连翘、花椒、刺梅等28 种树木。
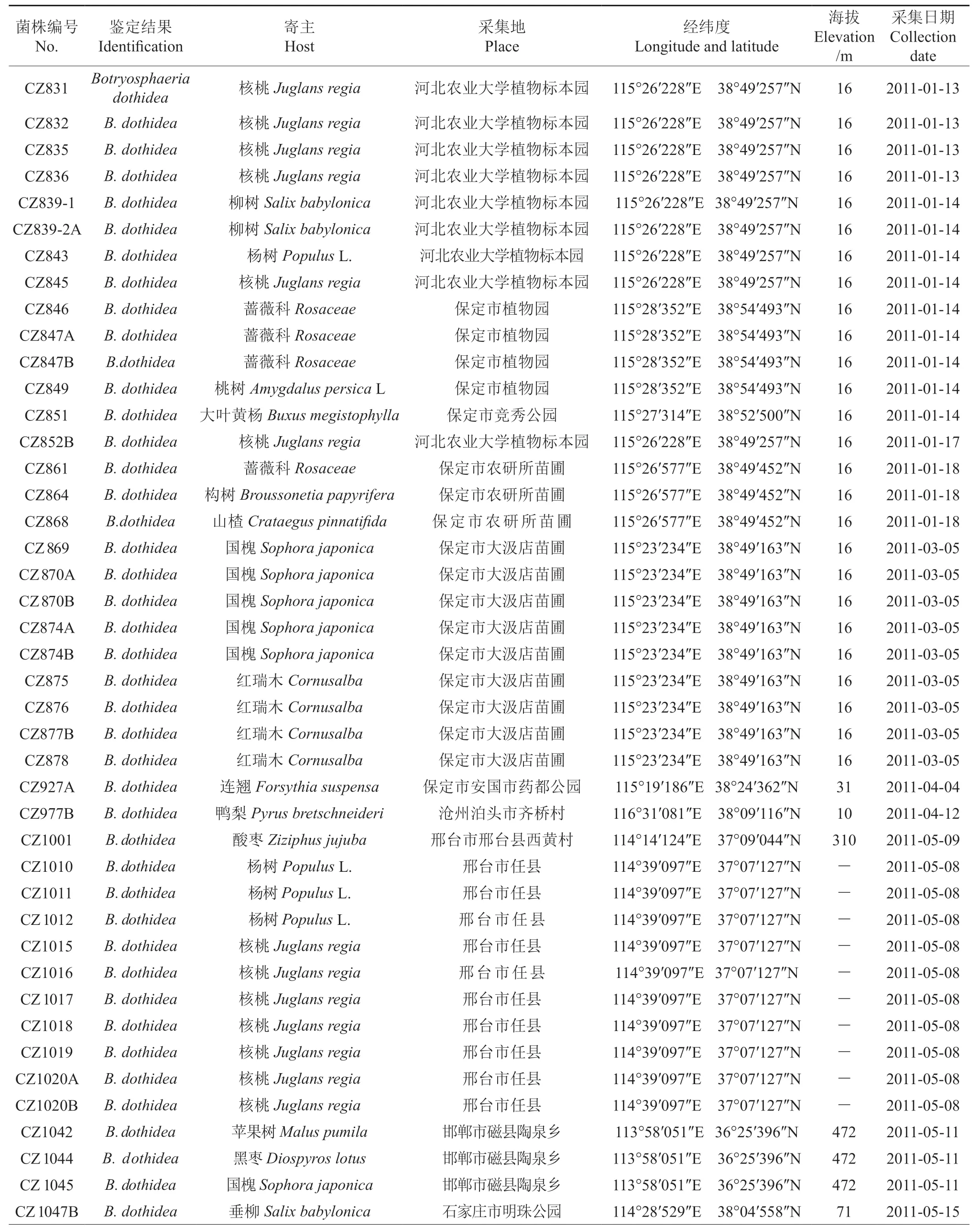
表1 本研究试验菌株的寄主植物,采集地点,采集地Table 1 The host plants, collecting locality of fungal isolates used in this study
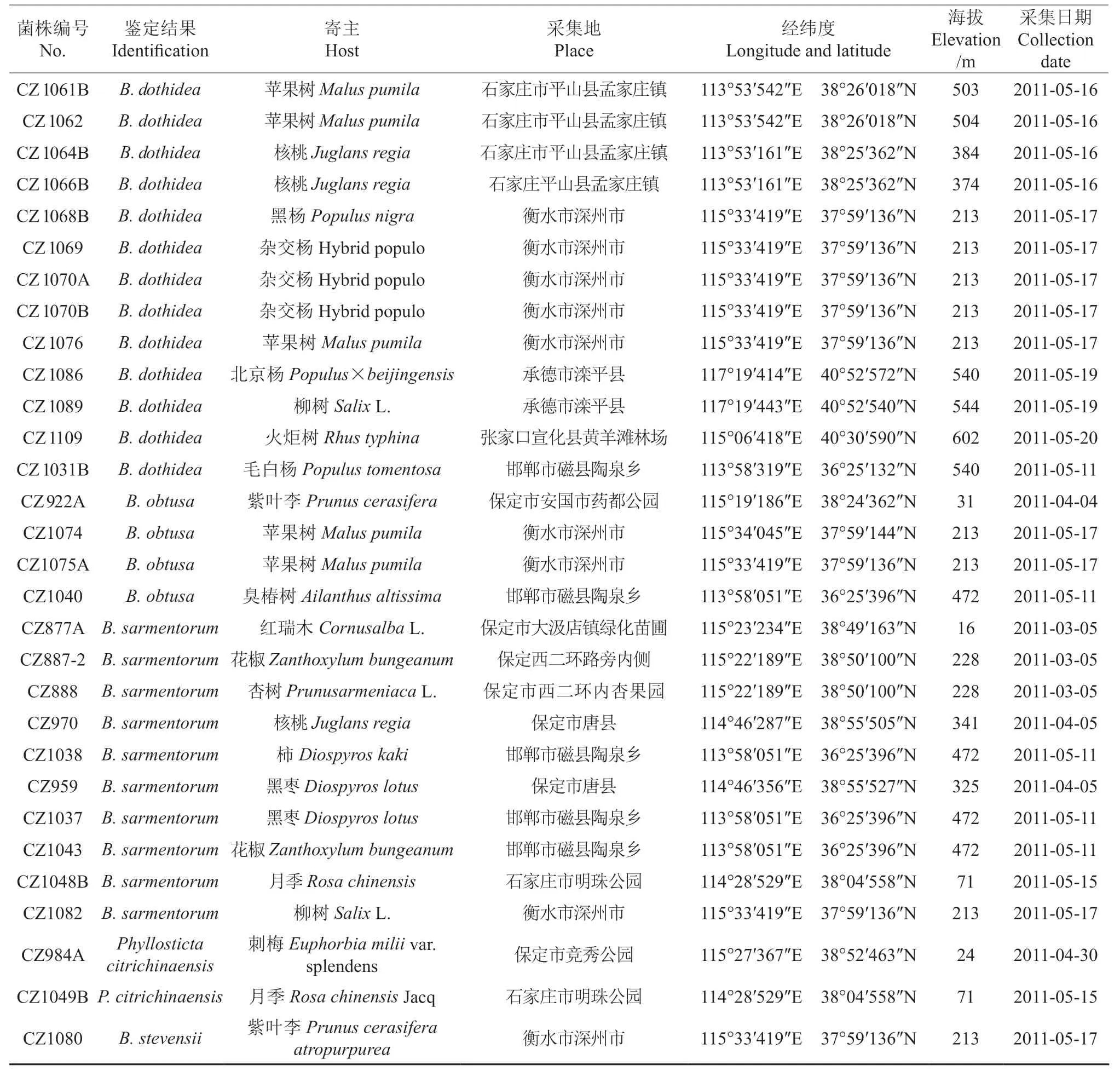
续表1Continuation of table 1
研究发现,所有试验菌株均能培养成单菌落,并且在载有西黄松针的水琼脂培养基上均可诱导形成理想的子实体。通过观察记录子实体形态特征和菌物菌落的培养学特征,将73 株试验菌株鉴定为5 种葡萄座腔菌科菌物:Botryosphaeria dothidea(共56 株)、B.obtusa(4 株)、B.stevensii(1 株)、B.sarmentorum(10 株)和Guignardia citricarpa(2 株)。5 种菌物的典型形态学及培养学特征分别描述如下:
Botryosphaeria dothidea的培养学特征(菌株CZ1066B):在PDA 培养基上生长4 d 后,菌物菌丝布满整个培养皿,比一般菌物生长快;菌丝最初为透明状,后逐渐变为白色、灰白色、至灰绿色,菌丝由疏松到致密,呈绒毛状。B.dothidea的形态学特征:分生孢子器为球形或器型、单腔室,表生,离散或聚生,分生孢子器直径0.31 ~0.46 mm。分生孢子器外壁由厚壁角质化组织构成,致密;分生孢子器中无分生孢子梗,有产分生孢子细胞,其从分生孢子器内壁层细胞上生出,全壁芽生,产分生孢子细胞无色,棒状;分生孢子为梭形,先端渐尖,基部截距状,薄壁,无色,无隔膜;分生孢子大小范围为(11.31 ~28.16)μm×(3.72 ~ 5.57)μm,50 个分生孢子的长、宽平均值及标准差 为(20.18±3.77)μm×(4.60±0.43)μm; 分生孢子长宽比(L/W)为2.78 ~6.58,50 个分生孢子的L/W 比的平均值及标准差为4.40±0.79(图1)。根据以上培养性状和形态特征,初步判定菌株CZ1066B 为菌种B.dothidea。
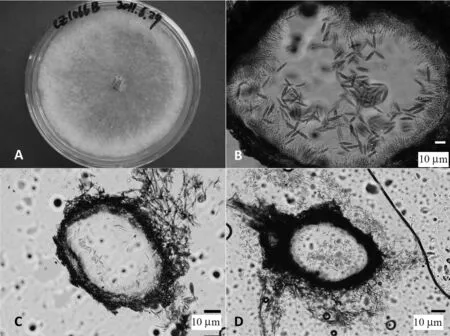
图1 试验菌株CZ1066B 的培养学及形态学特征Fig.1 Cultural and morphological features of the isolate CZ1066B
B.sarmentorum的 培 养 学 特 征( 菌 株CZ1038):菌株生长较慢,在PDA 培养基上生长4 d 后菌丝仍未长满培养皿(直径9 cm),菌落绿色,出现环状,致密,基本无气生菌丝体。B.sarmentorum的形态学特征:分生孢子器为球形或器型,单腔室至多腔室,单生、聚生或散生,灰黑色,直径范围为0.40 ~0.54 mm;分生孢子器棕黑色,由厚而致密的角质化组织构成;分生孢子圆柱形或卵圆形,顶部钝圆、基部截距状,浅棕色,半透明、光滑、无纹饰,具1 个膈膜;分生孢子大小范围为(13.19 ~19.34)μm×(5.76 ~9.66)μm, 50 个分生孢子的长、宽平均值及标准差为(16.61±1.58)μm×(7.88±0.84)μm;分生孢子长宽比(L/W)为1.68 ~2.85,50 个分生孢子的L/W 比平均值及标准差为2.12±0.25(图2)。根据以上培养学特征和形态学特征,初步判定菌株CZ1038 为菌种B.sarmentorum。
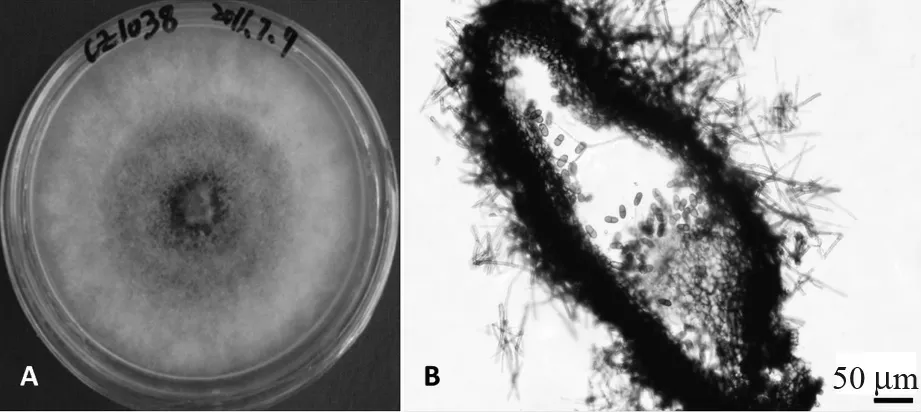
图2 试验菌株CZ1038 的培养学及形态学特征Fig.2 Cultural and morphological features of the isolate CZ1038
B.stevensii的培养学特征(菌株CZ1080):在PDA 培养基上6 d 菌丝未布满培养皿,比一般菌株生长较慢;培养初期为透明状菌落,后渐变为绿色及墨绿色,具疏松绒毛状菌丝。B.stevensii的形态学特征:分生孢子体器型至球形,单腔室,单生、聚生或散生,直径范围为0.34 ~0.52 mm; 分生孢子体壁厚,由致密的角质化细胞组成;分生孢子为卵形或圆柱形,顶端钝圆,深棕色,具1 个膈膜,光滑、无纹饰;分生孢子大小范围为(15.56 ~25.85)μm×(6.49 ~12.54)μm, 50 个分生孢子的长、宽平均值及标准差为(20.34±2.23)μm×(9.56±1.19)μm;分生孢子长宽比(L/W)为1.38 ~2.99,分生孢子的长宽比(L/W)平均值及标准差为2.15±0.31(图3)。由菌株的培养学特征和形态学特征可知,菌株CZ1080 为B.stevensii。
B.obtusa的培养学特征(菌株CZ1040):真菌在PDA 平皿上培养4 d 其菌丝就布满整个培养皿,与一般菌株相比生长较快;菌丝连续3 天透明,后变为灰色,疏松。B.obtusa的形态学特征:分生孢子体球形、器型或烧瓶状,单腔室至多腔室,单生、聚生或散生,直径范围为0.16 ~0.51 mm; 分生孢子体壁由致密的厚壁角质化组织构成;分生孢子圆柱形至卵形,端部圆,棕色,无膈膜,光滑,无纹饰;分生孢子大小范围为(14.07 ~ 23.15)μm×(6.95 ~10.15)μm,50 个分生孢子的长、宽平均值及标准差为(19.40±2.34)μm×(8.68±0.80)μm;分生孢子长宽比(L/W)为1.46 ~2.99,分生孢子的长宽比(L/W)平均值及标准差为2.25±0.33(图4)。根据以上培养性状和形态特征,菌株CZ1040 被初步判定为B.obtusa。
Phyllosticta citrichinaensis的培养学特征(菌株CZ984A):在PDA 培养基上生长6 d 菌丝还未铺满培养皿,比一般菌株生长较慢;菌丝由透明变为灰绿色,致密,呈粉末状。P.citrichinaensis的形态学特征:子囊呈棍棒状,包含8 个子囊孢子(子囊孢子不规则地分成两列)无色;子囊孢子卵圆形至椭圆形。子囊孢子大小范围为(5.1 ~ 9.83)μm×(4.96 ~7.91)μm,50 个 子 囊 孢 子的长、宽平均值及标准差为(8.01±1.02)μm×(6.18±0.66)μm;子囊孢子长宽比(L/W)为0.68 ~1.88,50 个孢子L/W 比的平均值及标准差为1.31±0.25(图5)。由菌株的培养学特征和形态学特征可知,菌株CZ984A 被初步判定为Guignardia citricarpa,是P.citrichinaensis的无性型。
2.2 rDNA-ITS 片段扩增及序列测定
利用引物对ITS1/LR5 扩增所有菌株的rDNAITS 序列片段,经过电泳检测,均能产生特异性良好的条带,基本上能产生单一且亮的条带,并且rDNA-ITS序列的测序结果理想。试验菌株的rDNA-ITS序列片段大小一般约为1 600 bp,然而菌株CZ984A、CZ1049B 的rDNA-ITS序列片段却较长,约为2 000 bp。部分试验菌株rDNA-ITS序列PCR 产物电泳结果见图6。
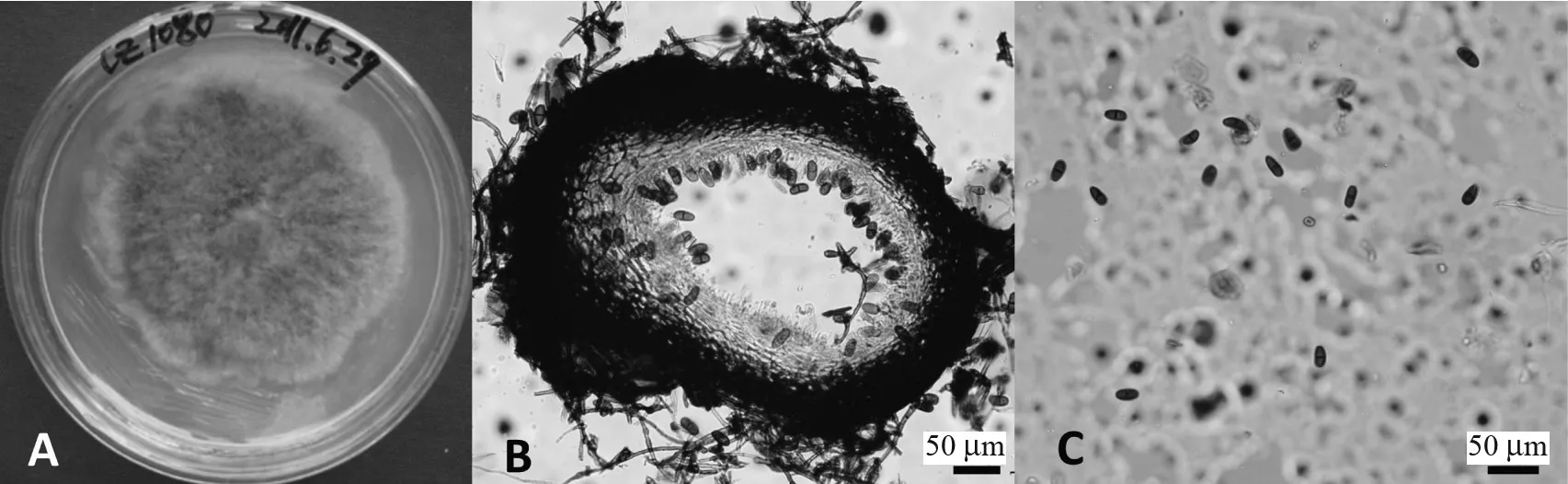
图3 试验菌株CZ1080 的培养学及形态学特征Fig.3 Cultural and morphological features of the isolate CZ1080
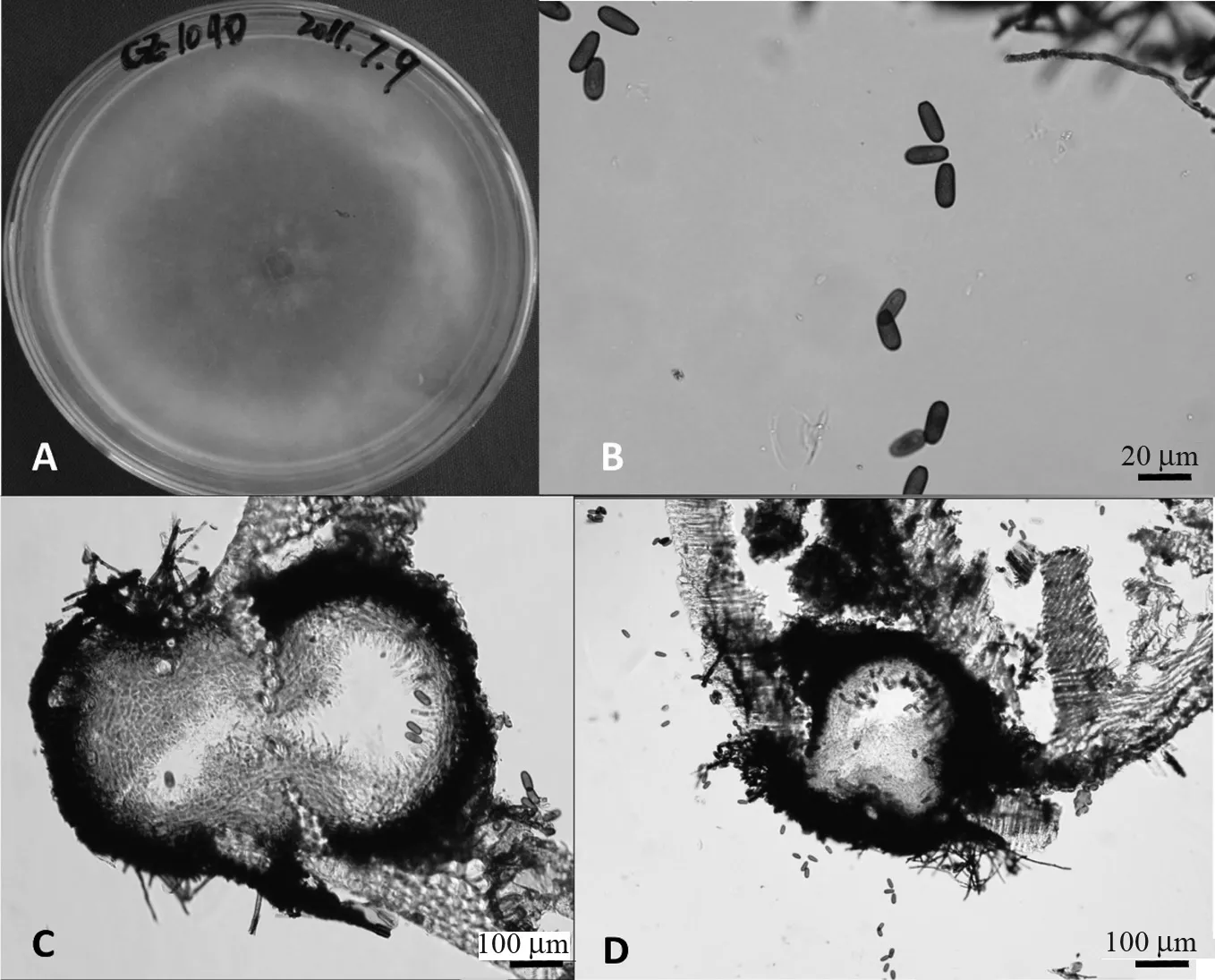
图4 试验菌株CZ1040 的培养学及形态学特征Fig.4 Cultural and morphological feature of the isolate CZ1040

图5 试验菌株CZ984A 的培养学及形态学特征Fig.5 Cultural and morphological features of the isolate CZ984A
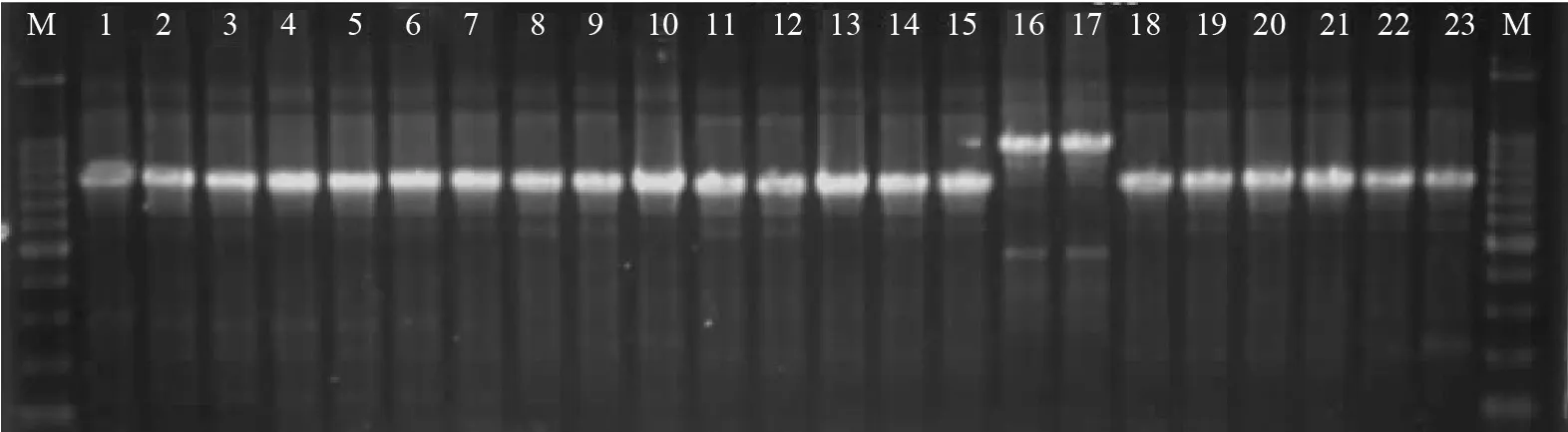
图6 部分试验菌株的rDNA-ITS序列扩增结果Fig.6 rDNA-ITS sequences amplication results of partial experimental isolates
2.3 rDNA-ITS序列的系统发育分析
以GenBank 数据库中的37 个葡萄座腔菌科真菌的rDNA-ITS序列作为参比序列,采用Kimura-2-parameter 遗传距离,以rDNA-ITS序列构建试验菌株的邻位连接法(Neighbor-joining,NJ)系统进化树。为了更清晰地揭示Botryosphaeria dothidea主要分支的系统发育关系,本研究选取河北省内不同地区且不同寄主的11 个B.dothidea代表菌株对原始进化树进行简化,结果见图7。
系统发育分析显示本研究的所有试验菌株均为葡萄座腔菌科菌物(图7)。以Kimura-2 距离构建的NJ 系统发育分析将共110 个试验及参比菌株聚类为5 个主要分支,分别对应于Lasiodiplodia/Diplodia、Phyllosticta、Fusicoccum、Neofusicoccum和Dothiodia这5 种 不 同LSU 类 群 的 无 性 型。Lasiodiplodia/Diplodia分支含有试验菌株:CZ1040、CZ922A、CZ1074、CZ1075A 与CZ1080;参 比 菌株:B.stevensii、B.obtusa、B.rhodina及其无性型菌株。在此分枝内形成一对姐妹群,即由参比菌株(B.obtusa、B.stevensii)与CZ1080、CZ922A、CZ1040、CZ1074、CZ1075A 构成的单系类群和由B.rhodina及其无性型菌株组成的单系类群。在这对姐妹群中,CZ1080 与B.stevensii亲缘关系较 近(BS=57),CZ922A、CZ1040、CZ1074、CZ1075A 与B.obtusa亲缘关系相近(BS=72)。利用分子鉴定和形态鉴定研究方法,最终将试验菌株CZ1080 鉴定为B.stevensii,将CZ1040、CZ922A、CZ1074、CZ1075A 这4 个 试 验 菌 株 鉴 定 为B.obtusa。Phyllosticta分 支 由2 个P.citrichinaensis菌株和2 个试验菌株CZ984A、CZ1049B 构成,且具有最高支持率(BS=100)。试验菌株CZ984A、CZ1049B 和参比菌株P.citrichinaensis的亲缘关系极其相近,结合形态学特征,最终将CZ984A、CZ1049B 鉴定为P.citrichinaensis。在Fusicoccum分支中,除了2 个B.dothidea外,其余都是CZ864、CZ876、CZ927A 等56 个试验菌株(BS=89),分子鉴定结合形态鉴定,将这56 个试验菌株最终鉴定为B.dothidea。在Neofusicoccum分支中只有参比菌株N.mediterraneum、N.australe、N.parvum、N.ribis和N.vitifusiforme,无试验菌株。在Dothiodia分支中,除了2 个B.iberica和2 个B.sarmentorum菌株外,其余都是CZ1038、CZ1043、CZ1048B、CZ1037、CZ959、CZ970、CZ888、CZ887-2、CZ877A 和CZ1082 等10 个试验菌株(BS=76),其中试验菌株聚成一小枝(BS=65);利用分子鉴定和形态鉴定研究方法,最终将这些试验菌株鉴定为B.sarmentorum。
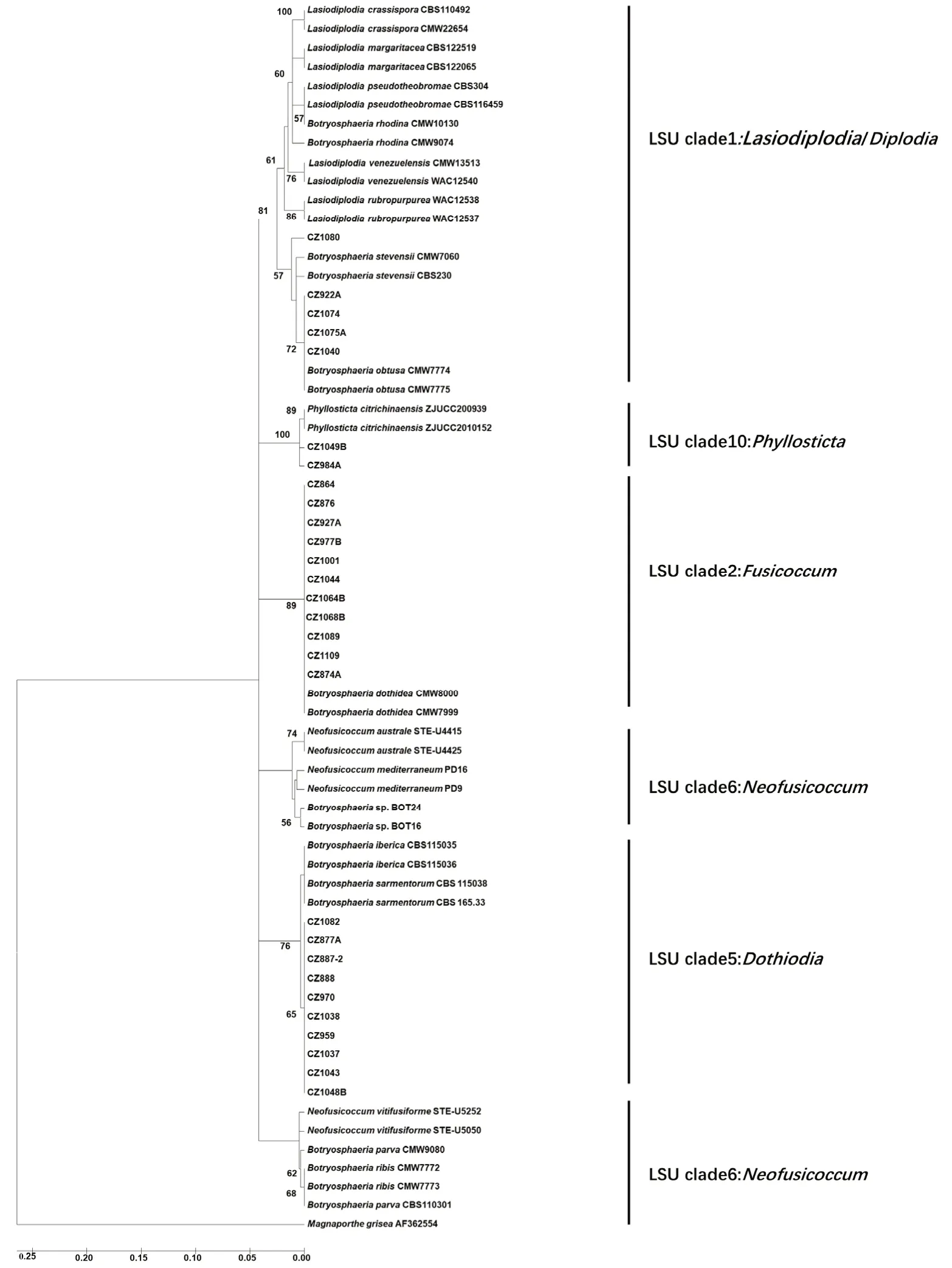
图7 简化后的rDNA-ITS 部分基因序列的Kimura-2-parameter NJ 系统发育树Fig.7 Kimura-2-parameter NJ curtness tree obtained from rDNA-ITS gene partial sequences data
以P 距离构建的NJ 系统发育分析同样将110 个试验菌株和参比菌株聚类为5 个主要的分支,分别对应于Dothiodia、Neofusicoccum、Lasiodiplodia/Diplodia、Fusicoccum和Phyllosticta等5 个不同的无性型(图8)。在Dothiodia分支中,除了2个B.sarmentorum和2 个B.iberica菌株外,其余都是本研究采集的10 个试验菌株(BS=86),而且这10 个试验菌株聚成一小枝(BS=64);根据形态鉴定和分子鉴定方法,最终将这些试验菌株确定为B.sarmentorum。在Neofusicoccum分支中只包含参比菌株N.ribis、N.australe、N.vitifusiforme、N.parvum和N.mediterraneum各2 个rDNA-ITS序列,但不包括试验菌株序列(BS=72)。Lasiodiplodia/Diplodia分 支 包 括B.obtusa、B.stevensii、B.rhodina及其无性型菌株,试验菌株CZ1040、CZ922A、CZ1074、CZ1075A和CZ1080(BS=85);在此分枝内形成一对姐妹 群:试 验 菌 株CZ1040、CZ922A、CZ1074、CZ1075A、CZ1080 与B.obtusa、B.stevensii聚成的单系类群和B.rhodina及其无性型菌株聚成的单系类群。CZ1080 与B.stevensii亲缘关系相近(BS=55),试验菌株CZ922A、CZ1040、CZ1074、CZ1075A 与B.obtusa菌株遗传距离较近(BS=81);结合培养学和形态学特征将试验菌株CZ1080 鉴定为B.stevensii菌种,将CZ1040、CZ922A、CZ1074、CZ1075A 这4 个 试 验 菌 株鉴定为B.obtusa菌株。在Fusicoccum分支中,除了2 个B.dothidea菌株外,其余都是试验菌株CZ864、CZ876、CZ927A 等56 个(BS=100),结合培养学和形态学特征将这些试验菌株最终鉴定为B.dothidea。Phyllosticta分支仅包括2个Phyllosticta citrichinaensis菌 物 和CZ984A、CZ1049B 等2 个试验菌株(BS=100),结合培养学和形态学特征将这2 个试验菌株最终鉴定为P.citrichinaensis。
3 结论与讨论
有些国内专家学者相继统计或者研究报道葡萄座腔菌科菌物的寄主[1,22-26],2015年姜晓龙等首次报道浸染蓖麻Ricinus communisL.的病原菌物为B.dothidea[27],2018年朱琪丽等首次报道葡萄座腔菌侵染柑橘Citrus reticulataBlanco.果实[28],2018年黄静等发现落叶松葡萄座腔菌引起草莓Fragaria ananassaDuch.病害[29]。Chen 等在希腊和美国加利福尼亚的开心果Pistacia veraL.中发现2 种新的葡萄座腔菌科菌物Neofusicoccum hellenicum sp.nov.和Lasiodiplodia americana sp.nov.[30]。Aćimović 等 利 用ITS 和EF-1α序 列 分析,鉴定加利福尼亚海岸红杉Sequoia的病原菌物为B.dothidea[31]。本研究根据大量的相关资料,发现6 个种寄主植物,即大叶黄杨Buxus megistophylla、 构 树Broussonetia papyrifera、 火炬树Rhus typhina、连翘Forsythia suspensa、花椒Zanthoxylum bungeanum和刺梅Euphorbia milii var.splendens为葡萄座腔菌科菌物的新寄主植物。
为了明确引发河北省树木溃疡病的葡萄座腔菌科菌物种类,本研究采用无性型形态学和rDNA-ITS序列对该类菌物进行系统分类研究,鉴 定 出B.dothidea、B.obtusa、B.stevensii、B.sarmentorum和Phyllosticta citrichinaensis等5 种葡萄座腔菌科菌物。B.dothidea是一类常见树木溃疡病病原菌物,分布广泛、寄主多样,其无性型为七叶树壳梭孢Fusicoccum aesculi[32]。本研究将73 个试验菌株鉴定为葡萄座腔菌科菌物,其中56 个为B.dothidea菌株,占试验菌株总数的77%;这些B.dothidea菌株分别侵染核桃、苹果、桃树、梨树、黑杨、北京杨、柳树、国槐、山楂、连翘等20 种植物(甚至包括了外来入侵树种火炬树),显示出该种菌物的寄主广泛性。CZ1038、CZ1043、CZ1048B、CZ1037、CZ959、CZ970、CZ888、CZ887-2、CZ877A 和CZ1082 等10 个试验菌株具有Dothiorella型分生孢子,被鉴定 为B.sarmentorum。2005年Phillips 等 曾 报 道B.sarmentorum和B.iberica这2 种病原的形态学特征[33]。在Neofusicoccum分支内,B.parva真菌无性型Neofusicoccum parvum是引发全球多种植物病害的病原[34]。Neofusicoccum ribis是B.ribis真菌无性型名称[35],B.ribis与B.parva向来很难区分,2009年Pavlic 等采用多基因序列分析,明确B.parva和B.ribis是2 个不同的生物学种[36]。在Lasiodiplodia/Diplodia分支内,试验菌株CZ922A、CZ1040、CZ1074、CZ1075A 和CZ1080均具有Diplodia型分生孢子,最终判定CZ1080 为菌种B.stevensii,其他4 个试验菌株为B.obtusa。B.stevensii、B.obtusa的无性型分别是Diplodia mutila、D.seriata。D.mutila引发杨树溃疡病害已经有很多报道,D.seriara则在2011年被首次报道为我国杨树溃疡病的病原[23]。在Phyllosticta分支中,菌株CZ984A 和CZ1049B 作为Phyllosticta属,明显和Botryosphaeria属分为2 大分支。Phyllosticta属和葡萄座腔菌属的不同之处:一是试验菌株CZ984A 和CZ1049B 的DNA 片段比其他试验菌株的DNA 片段较长;二是通过rDNAITS 序 列 同 源 比 对,CZ984A 和CZ1049B 没 有ITS4 序列,其ITS1 序列上游比其他菌株多出一段DNA 片段。
目前,在生物系统分类研究中,很多与生物体基本功能相关且具有结构保守性的基因或者序列被广泛用于揭示生物之间的系统发育关系,如在植物学中的叶绿体ndhF 和rbcL[37]。β-tubulin 基因和EF-1α基因[18-19,21,30]等广泛应用于菌物的系统发育研究,但是ITS-rDNA 序列仍然是植物和菌物系统分类中应用最广泛的基因片段[10,12,16,18-19,30,38-40]。李文英等指出,以物种全型的培养特性、形态特征及多基因序列佐证等作为重要分类依据,是葡萄座腔菌科菌物类群分类学和系统发育研究的必然趋势[41]。本研究采用ITS-rDNA 序列,同时用Blast 方法搜集全部NCBI 中的相关菌株的基因序列,应用聚类分析方法对真菌进行分类;在此基础上,结合真菌培养学性状和形态学特征对河北省枝干溃疡病菌进行分类鉴定,结果具有较高的可靠性。
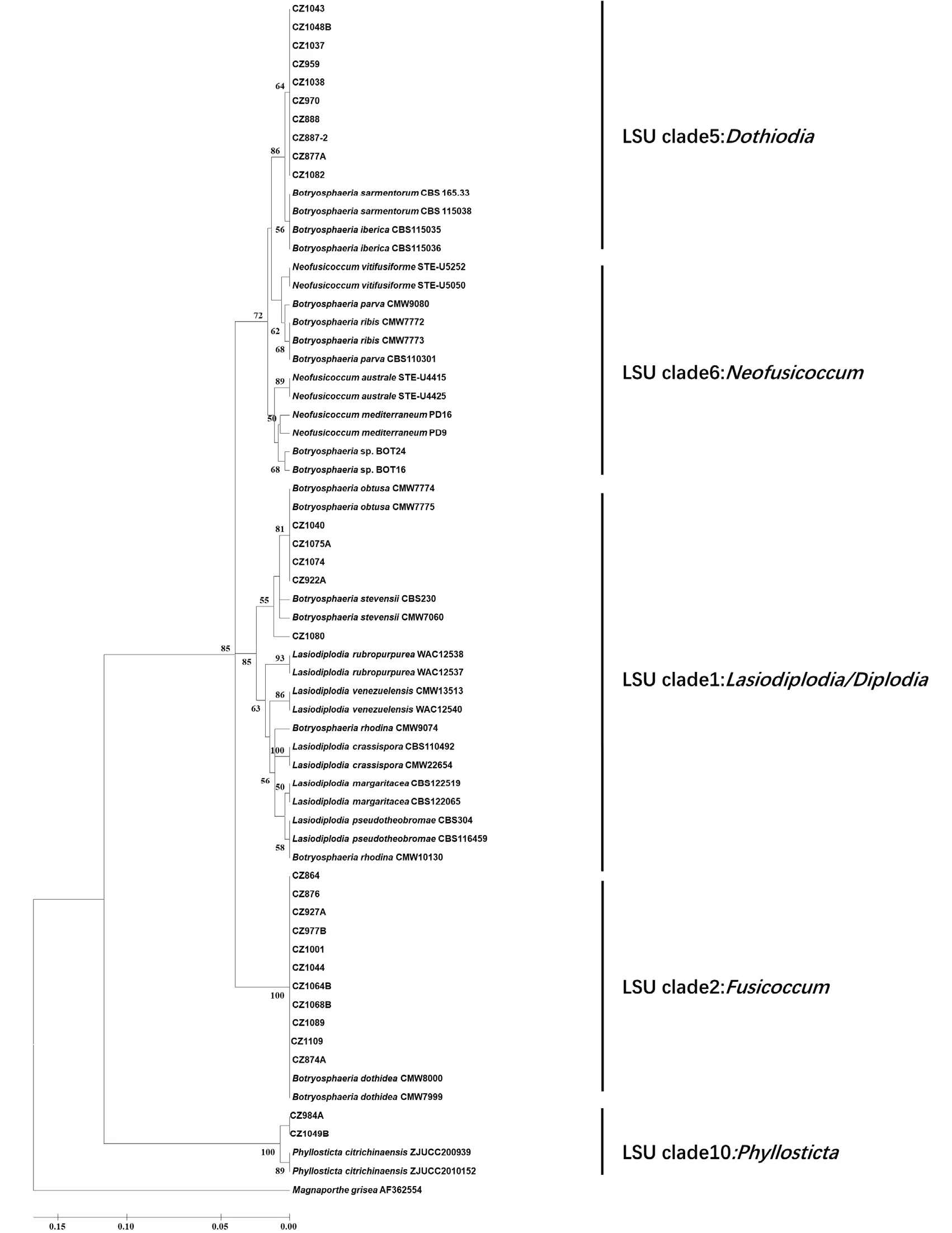
图8 简化后的rDNA-ITS 部分基因序列的P-距离NJ 系统发育树Fig.8 P-distance NJ curtness tree obtained from rDNA-ITS gene partial sequences data
本研究首次对河北省树木溃疡类病害病原菌物分布及系统分类的分析,初步明确了该地区溃疡类病害的寄主植物范围并发现6 种新的寄主植物,为河北省树木溃疡病的防治和树木溃疡病菌物的系统分类研究提供理论依据。研究结果显示,葡萄座腔菌B.dothidea仍然是危害河北省树木健康生长的最为重要、分布最为广泛的枝干病原菌物,不仅危害杨树、核桃、苹果等传统寄主,而且危害到花椒等新寄主植物,甚至是入侵树种火炬树。因此,有必要对B.dothidea溃疡致病性分化和种群遗传分化进行深入分析,从而提出河北省以及邻近地区树木溃疡类病害的防治策略。本研究的系统发育分析显示,CZ1038、CZ1043 等10 个菌株以较高支持率(p 距离进化树:BS=64;Kimura-2 距离进化树:BS=65)单独聚为一个分枝,然后与B.sarmentorum和B.iberica菌株聚类形成LSUDothiodia聚类群(图7 和图8),该类群的菌株具有Dothiorella型分生孢子。由于没有观察到与B.sarmentorum可区分的形态学特征,这些菌株被初步确定为B.sarmentorum,然而ITSrDNA 聚类分析结果提示我们这些菌株可能构成一个新种。因此,未来需要在形态学特征观察的基础上,采用多基因分类的方法以最终确定这些菌株的系统分类地位。
