食用菌内源脂肪酸的研究
2020-02-12张梦洁
张梦洁
(江苏省建筑工程质量检测中心有限公司,江苏 南京 210000)
不饱和脂肪酸,特别是长链多不饱和脂肪酸,是维持健康必要物质,其对于降低血液胆固醇及调节细胞生理具有重要作用[1-2]。膳食中含有的必需脂肪酸如亚油酸和α-亚麻酸等[3],人体自身却无法自行合成[4-5]。然而,无论植物性还是动物性油脂都含有较高热量,由于高脂类食品对健康不利,因此急需寻求健康脂肪酸来源。在众多食物中,食用菌既可提供必需脂肪酸,还具有低卡路里、无胆固醇、高纤维和高蛋白质等优点[6]。因此,与脂肪来源食物花生、大豆、肉类和其他常见蛋白质相比,食用菌营养价值更为显著。
在众多的不饱和脂肪酸中,棕榈酸(16:0),油酸(18:1) 和亚油酸(18:2) 是担子菌门(Basidiomycota) 中最常见的不饱和脂肪酸[7-8]。有研究在双孢菇(Agaricus bisporus)、草原黑蘑(Agaricus arvensis)、赭盖鹅膏菌(Amanita rubescens)、姬松茸(Agaricus blazei)、牛肝菌(Boletus subglabripes)、鸡腿菇(Coprinus comatus)、灵芝(Ganoderma lucidum) 和马勃(Lycoperdon pyriforme) 中鉴定到16:1,Δ9;16:1,Δ11;17:1,Δ9;18:1,Δ9;18:2,Δ9、12;20:1,Δ11;20:2,Δ11、14;22:1,Δ13 和24: 1,Δ15 等不饱和脂肪酸[8-10]。此外,研究发现同一种食用菌的生长环境影响其脂肪酸浓度和类型[11]。食用菌中蛋白质含量也较高,甚至含有比牛肉更优的氨基酸组成比例[12]。此外,研究发现食用菌具有消炎[13-14],抗菌[15],预防糖尿病[16-17]等药理活性及抗氧化活性[18]。香菇多糖体具有加强人体免疫能力和抑制肿瘤细胞生成的作用[19]。有研究表明,在肿瘤病人化疗期间应用浓缩“多糖体”,对病人具有显著治疗效果[19]。因此,通过对食用菌营养成分中脂肪酸研究进展进行概述,可为开发利用食用菌提供参考。
1 食用菌特性与栽培
约有220 万种到380 万种真菌分别归属不同形态特征门类[20],分为多细胞(丝状真菌和食用菌)和单细胞(酵母) 真核微生物[21]。担子菌门(Basidiomycota) 是Hibbett 等[22]提出的7 个真菌门之一。据统计,担子菌门大约有140 000 种食用菌,其中只有10%~15%具有分类学特征[23-24]。食用菌形态特征较简单,由菌盖、菌柄、菌托、菌环等部分组成[25]。菌肉存在于皮层下方,由长型丝状或膨大泡囊状菌丝组成。菌肉及受损菌肉颜色常因种类不同存在差异[26]。食用菌鲜品主要由水(约90.0%),碳水化合物(2.5%~5.8%),蛋白质(2.6%~4.0%),脂肪(0.2%~0.7%) 和纤维(0.6%~1.1%) 组成[27-28]。
相对于其他农作物,食用菌生产对栽培场所及条件要求较低,如草菇(Volvariella volvacea)、金针菇(Flammulina velutipes)、平菇(Pleurotus ostreatus) 和杏鲍菇(Pleurotus eryngii) 等已在世界各地广泛栽培。根据联合国粮食及农业组织的数据报告,2017 年世界上生产的食用菌总量达1 020 万吨,2007 年至2017 年期间,亚洲食用菌产量占全球产量76.0%,其次是欧洲(17.2%)、美洲(5.9%)、大洋洲(0.6%) 和非洲(0.2%)[34]。但食用菌生产也需特定条件控制,如恒湿、适宜温度、通风、光照和培养底物[29-30]等,这些环境条件变化能直接影响食用菌形态和营养价值。如平菇在温和条件比高温条件长得更好[31]。也有研究发现,对于同一食用菌物种,菌丝在24℃~32℃环境温度下最适生长[32]。
营养风味方面,Patil 等[33]研究了不同基质上生长的平菇蛋白质、钙和铁含量变化,结果显示平菇在大豆废弃物中培养,营养物质含量最高。另外,食用菌不同部位营养成分含量差异明显,如红菇(Russula griseocarnosa) 菌盖部分富含镁、磷、钠、脂质和蛋白质,而菌柄部分具有更高的铁、钙和纤维含量[34]。Kalac 比较了自然生长和人工栽培食用菌的营养价值,发现同一物种在不同生长条件下其营养成分存在明显差异,如金针菇(Flammulina velutipes) 干物质中,游离氨基酸含量的差异为1.5 g·kg-1~72.0 g·kg-1[35]。
尽管已有较多文献详细报道食用菌的营养价值,但都集中于人工栽培食用菌品种,还有多种野生食用菌营养价值仍未得到充分研究[36]。野生食用菌开发较缓慢的原因主要有两方面,一是人工栽培技术还未能模拟野生食用菌生长环境,二是某些野生食用菌具有潜在毒性。在云南,已知可食用菌有800多种,剧毒野生菌近500 种[37]。目前,还未找到明显形态特征以区分食用和有毒食用菌。即便在一个属中,也有可食用和剧毒性物种[38-39],如红菇属可食用的冷杉红菇(Russula abietina) 和有毒的诱吐红菇(Russula emetica)。因此,食用菌中毒屡见不鲜,每年都会有严重中毒事件报道[40]。
2 脂肪酸种类及其营养价值
脂类物质是重要能源物质,也是必需脂肪酸的来源,同时还是生物体重要组成成分,而脂肪酸则是脂类的关键成分[1]。脂肪酸结构简单,自然界中约有40 多种脂肪酸,通常由2 碳单位生物合成,因此其中含偶数碳原子的种类较多[1]。脂肪酸除根据碳链长度不同进行分类外,还可根据饱和度情况及营养价值分类[41-42。其中,必需脂肪酸人类无法自身合成,只能依赖食物等外界供给,均属于ω-3 族和ω-6 族多不饱和脂肪酸[43]。
脂肪酸命名以碳链中碳原子数:双键数表示。可通过2 种方式计算碳原子数:羧基碳原子为1,双键位置碳原子距碳原子1 之间碳原子数以Δ(δ)上标表示,如油酸(18:1,Δ9) 表示含18 个碳原子,一个双键,在第9 位~第10 位碳原子间有一双键;或距离羧基最远的甲基碳原子称为ω-碳,其位置标为1 开始计数,按字母编号依次为ω-1、ω-2、ω-3等。如亚麻酸(18:1, ω-3),表示含有18 个碳原子,1 个双键,在C-3 和C-4 之间具有双键[44]。
哺乳动物能合成只含1 个双键的不饱和脂肪酸。如能通过去饱和酶(Δ9) 将饱和硬脂酸(C18:0) 转化为单不饱和油酸(C18:1)[45]。某些动物能通过其他去饱和酶(如Δ5 和Δ6) 生成多不饱和脂肪酸ω-6(花生四烯酸)、ω-9(正二十碳五烯酸) 和二十二碳六烯酸(DHA)。亚油酸(C18:2) 和亚麻酸(C18:3) 也能转化为具有20 个和22 个碳原子的长链脂肪酸。由于缺乏ω-3 去饱和酶,人类不能合成含有2个或2 个以上双键的多双键脂肪酸(如亚油酸和亚麻酸) 即必需脂肪酸,其必须从食物中获取[1]。必需脂肪酸参与到人体高密度脂蛋白形成,该脂蛋白能降低罹患心血管疾病的风险[1]。此外,ω-3 脂肪酸能影响细胞膜结构、细胞蛋白功能、脂质介质产生和基因表达模式,从而改善人类健康状况[46-47]。但必需脂肪酸需要保持均衡摄入,如摄入ω-6 ∶ω-3 比例为1 ∶1 或2 ∶1 时可预防肥胖,而不平衡的ω-6 ∶ω-3将影响脂肪合成。
食用菌含有丰富的饱和脂肪酸和不饱和脂肪酸,其中常见的饱和脂肪酸主要是软脂酸(C16) 和硬脂酸(C18),而不饱和脂肪酸主要是棕榈酸(C16:1,Δ9)、油酸(C18:1,Δ9)、亚油酸(C18:2,Δ9、12)、亚麻酸(C18:3,Δ9、12、15)、二十二碳六烯酸(C22:6, Δ4,7,10,13,16,19) 和二十碳五烯酸(C20:5,Δ5、8、11、14、17)见图1。
3 食用菌脂肪酸含量
比较食用双孢菇(Agaricus bisporus) 与其他食品的脂肪酸模式见表1,对不同种类食用菌或食用菌不同部位进行脂肪酸分析见表2。
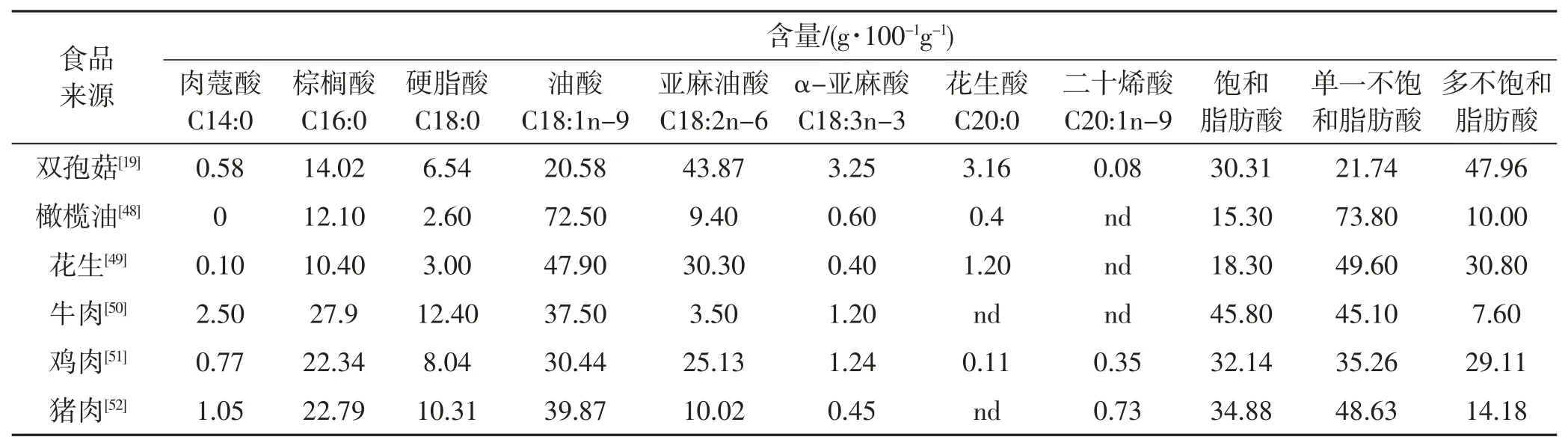
表1 部分常见食品的脂肪酸含量Tab.1 Content of fatty acids in common foods

表2 食用蘑菇中常见的不饱和脂肪酸的含量Tab.2 Content of common fatty acids in mushrooms
由表1、表2 可知,食用菌脂类物质含较高含量单不饱和脂肪酸和多不饱和脂肪酸,但饱和脂肪酸含量远低于肉类食品。在Ayaz 等[56]报道中,草原黑蘑和牛肝菌亚油酸含量分别达78.0%和75.6%,管形喇叭菌油酸含量达57.8%。李谭瑶等[6]利用气相色谱-质谱(GC-MS) 法对对市售的竹荪(Dictyophora indusiata)、双 孢 菇(Agaricus bisporus)、 香 菇(Lentinus edodes)、杏鲍菇(Pleurotus eryngii)、茶树菇(Agrocybe aegerita)、金针菇(Flammulina velutipes)、黑木耳(Auricularia auricula) 中脂肪酸进行检测,共检测到37 种脂肪酸,其中亚油酸在6 种蘑菇中含量最高,竹荪中人体必需多不饱和脂肪酸含量最高且种类最多。Dogan 和Akbas[65]在橙盖鹅膏菌(Amanita caesarea) 中鉴定到37 种脂肪酸,其中油酸含量最高(58.0%),其次是亚油酸(19.0%)、软脂酸(15.0%) 和硬脂酸(6.1%)。Yilmaz 等[53]检测了双孢菇(Agaricus bisporus)、Agaricus campestris,白牛肝菌(Boletus edulis)、毛鬼伞菌(Coprinus comatus)、平菇(Pleurotus ostreatus)、长根小奥德蘑(Oudemansiella radicata) 和蜜环菌(Armillaria mellea) 共7 种食用菌的脂肪酸含量,结果发现亚油酸是其主要脂肪酸,其次是软脂酸、油酸、硬脂酸和花生四唏酸;对菌盖和菌柄脂肪酸含量检测发现食用菌脂肪酸主要分布于菌盖,菌盖:菌柄含量约为18:2。据此推测菌盖营养价值比菌柄高。通过借助高效薄层色谱(HP-TLC) 和GC-MS 技术,Hanus等[66]分析了15 种牛肝菌属菌类脂肪酸成分,结果显示该属菌类主要脂肪酸为亚油酸(C9,12-18:2,38%~58%)、油酸(C9~18:1,15%~42%) 和软脂酸(C16:0,7%~17%)。Marekov 等[67]对9 个属中的15 种食用菌进行营养成分分析,结果显示亚麻酸、油酸、软脂酸和硬脂酸占总脂肪酸的83.7%。在黄柄笼头菌(Endoptychum agaricoides) 中,亚麻酸的含量为62.9%,而变黑腊伞菌(Hygrocybe nigrescens) 主要是油酸,为42.7%。此外,在11 种菌类中还检测到少量的α-亚麻酸及其异构体物质[64]。Pereir 等[68]也报道了类似结果,亚油酸是黄鹅膏菌(Leucoagaricus leucothites) 的主要脂肪酸成分,油酸是粉褶白环蘑(Amanita umbrinolutea) 的主要脂肪酸成分,而夏生灰球菌(Bovista aestivales) 和深凹杯伞菌(Clitocybe gibba) 脂肪酸主要成分分别是软脂酸和硬脂酸。Kayode 等[62]对凤尾菇(Pleurotus sajor-caju) 脂肪酸图谱分析发现单不饱和脂肪酸和多不饱和脂肪酸的含量分别占总脂肪酸的49.0%和41.0%,其中油酸、亚油酸和亚麻酸的含量分别是41.7%、29.5%和11.6%,而饱和脂肪仅占9.6%。Obodai 等[69]在4 种栽培菌翘鳞香菇(Lentinus squarrosulusstrain LSF 和Lentinus squarrosulusstrain SQW)、 平菇(Pleurotus ostreatus) 和黑木耳(Auricularia auricula) 和2 种野生菌虎奶菇(Pleurotus tuberregium) 和粗柄白蚁伞(Termitomyces robustus)中检测到20 多种脂肪酸。Woldegiorgis 等[70]利用气相色谱-火焰离子化检测仪分析6 种野生食用菌类(Agaricus campestris、Laetiporus sulphureus、Termitomyces clypeatus、Termitomyces microcarpus、Termitomyces letestui、Termitomycesspp.) 和3 种栽培菌类(Pleurotus ostreatus、Laetiporus edodes、Agaricus bisporus) 脂肪酸图谱,显示亚油酸是主要的脂肪酸成分(32.7%~70.5%),其次是油酸(2.5%~27.8%)、软脂酸(11.4%~19.3%) 和硬脂酸(0.8%~6.7%),同时还检测到少量的必须脂肪酸α-亚麻酸和γ-亚麻酸,在所有物种中均检测到奇数碳原子的脂肪酸(C15:0) 和(C17:0)。奇数碳原子的脂肪酸在真菌中很少见,可能在合成过程中整合了不同的起始单元,或是偶数碳原子脂肪酸缺失一个碳原子所致。蚁巢伞属(Termitomyces) 的小白蚁伞菌(Termitomyces microcarpus) 含有丰富的软脂酸(27.2%)、γ-亚麻酸(19.7%) 和棕榈酸(12.8%),同时还检测到少量的α-亚麻酸、油酸、硬脂酸、二十二酸六烯酸(DHA)、亚麻酸、肉豆蔻酸和异油酸[71]。在蚁巢伞属的其他物种中,如刺状鸡纵菌(Termitomycessp.)和球盖白蚁伞(Termitomyces globulus) 中,研究人员检测到较高的DHA,分别为19.5%和33.0%[72]。DHA 对于人脑发育具有重要作用[70]。因此,富含DHA 的食用菌营养价值更高。
综上,食用菌总脂肪酸以不饱和脂肪酸为主,以亚油酸和油酸为主要成分[62,73],某些种类存在高含量的棕榈酸和硬脂酸。上述研究中食用菌中亚麻酸的含量较低,但也有研究发现印度地区食用菌白齿菌(Hydnum repandum) 和高大环柄菇(Macrolepiota procera) 以及巴基斯坦地区的大肥菇(Agaricus bitorquis) 中亚麻酸的含量较高[55,57]可能是由于不同地区的气候和土壤特性所致。
4 展望
食用菌中的脂肪酸种类丰富,且主要是不饱和脂肪酸,约有70.0%是亚油酸、亚麻酸和油酸。尽管不同种类的食用菌中脂肪酸的含量有差异,但都可作为人类膳食中必需脂肪酸来源。食用菌作为世界卫生组织和粮农组织推荐的“一荤一素一菌”的三大人类健康膳食结构的组成部分,食用菌消费量也随之日益增长。食用菌的生产方式由人工栽培向机械化生产发展,在世界多地区已基本实现食用菌工厂化栽培。通过了解不饱和脂肪酸的合成代谢路径,研究食用菌脂肪酸分子调控机制可为为食用菌育种及其脂肪酸利用提供理论依据。
