Larix olgensis growth-climate response between lower and upper elevation limits:an intensive study along the eastern slope of the Changbai Mountains,northeastern China
2020-01-18JianYuQijingLiu
Jian Yu·Qijing Liu
Abstract Larix olgensis is a dominant tree species in the forest ecosystems of the Changbai Mountains of northeast China.To assess the growth response of this species to global climate change,we developed three tree-ring width and biomass chronologies across a range of elevations in the subalpine forests on the eastern slope of the Changbai Mountains.We used dendroclimatic analyses to study key factors limiting radial growth in L.olgensis and its variation with elevation. The statistical characteristics of chronologies suggested that elevation is a determinant of tree growth patterns in the study area.Response function analysis of chronologies with climate factors indicated that climate-growth relationships changed with increasing elevation:tree growth at high elevation was strongly limited by June temperatures of the previous year,and as elevation decreases,the importance of temperature decreased;tree radial growth at mid-elevation was mainly controlled by precipitation towards the end of the growing season of the current year.Biomass chronologies reflected a stronger climatic signal than tree-ring width chronologies.Spatial correlation with gridded climate data revealed that our chronologies contained a strong regional temperature signal for northeast China.Trees growing below timberline appeared to be more sensitive to climate,thus optimal sites for examining growth trends as a function of climate variation are considered to be just below timberline.Our study objective was to provide information for more accurate prediction of the growth response of L.olgensis to future climate change on the eastern slope of the Changbai Mountains,and to provide information for future climate reconstructions using this tree species in humid and semi humid regions.
Keywords Tree-ring width·Dendrochronology·Changbai Mountains·Elevation gradient·Timberline·Larix olgensis
Introduction
The effects of global warming in northeastern China have become increasingly apparent over the past century(Wang and Liu 2011;Wang et al.2015).Over this period climate has been identified as the strongest driver of tree and forest growth rates(exceeding soil and anthropogenic disturbance)(Toledo et al.2011).As the strongest relationship between climate change and radial growth exists at the distribution limit of a species(Liang et al.2014),trees growing at the boundary of their species range are considered to be the most sensitive and vulnerable to future climate change.In the subalpine/alpine zone,tree radial growth is subject to environmental gradients associated with elevation(Gou et al.2005;Zhao et al.2014;Kharal et al.2017).An understanding of tree growth over elevation gradients,using the approach of space-for-time substitution,can therefore be used to evaluate and predict forest variations in response to future climate change.
Due to their high levels of information content regarding tree growth characteristics(e.g.,accurate age-dating,high resolution,wide distribution,accessible sites and long time series),tree-rings have widely been used to examine relationships between climatic factors and tree growth(Wang et al.2013;Fang et al.2015).Tree-ring analysis has also been widely used in palaeoclimate reconstructions(Davi et al.2015;D'Arrigo et al.2015).Shi et al.(2012)and Gao et al.(2013)documented the effects of climate on tree-ring width along elevation gradients using dendrochronological techniques.Climatic factors limiting tree radial growth at the upper timberline ecotones can be considerably different from those at lower elevations in mountainous areas(Liu et al.2013;Shen et al.2015).Generally,tree growth at the upper tree line is limited by low temperatures,while at lower elevations water availability is often the limiting factor(Lo et al.2010;Dang et al.2013;Sidor et al.2015).Liang et al.(2014),however,highlighted that the growth of Betula utilis at upper timberlines is typically limited by moisture availability,i.e.,a drought-induced alpine timberline.Other studies have recorded different results that challenge this general principle,with uniform growth patterns being observed at both the upper and the lower forest limits in different regions(Liu et al.2013;Yang et al.2013).As elevation has also been shown to have no significant effect on growth-climate relationships(Wang et al.2005; He et al. 2013), the physiological mechanisms determining tree growth response to climatic parameters along an elevation gradient are complicated. Detailed understanding of tree growth over elevation gradients is therefore essential to evaluate and predict variations in the potential response of forests to future climate changes in mountainous regions.
The Changbai Mountains are considered to be sensitive to global climate change(Lin et al.2013).This area has increasingly been the focus of dendrochronological studies(Yu et al.2007,2011;Wang et al.2013;Zhuang et al.2017).L.olgensis forests are distributed across a wide elevation range in the Changbai Mountains from 500 to 1950 m a.s.l on northern slopes,and this species often dominates local forest ecosystems. Knowledge of the dendrochronological features of this species across elevation gradients is important to understand and protect the L.olgensis forest ecosystem(Chen et al.2011;Shen et al.2015). Previous studies have investigated tree growth variations of L.olgensis in response to climate regimes at different elevations in this area,especially on the northern slope(Yu et al.2005b;Chen et al.2011;Shen et al.2015).Results from previous studies have indicated that tree growth response to climate change is complicated by differences in topography and climate,and that there is a need to investigate growth-climate relationships in different areas with varying climates. The eastern slope of the Changbai Mountains has unique topographic features and natural conditions,thus by investigating how elevation influences tree growth in this area, we can test the hypothesis that climate and radial growth relationships vary along the elevation gradient on this mountain.
The specific objectives of this study were:(1)to develop tree-ring width and biomass chronologies using data from L.olgensis along an elevation gradient on the east slope of Changbai Mountain;(2)to quantify the most important climatic factors influencing radial growth of L.olgensis with increasing elevation;(3)to understand the growth process of L.olgensis at different elevations;and(4)to explore differences in sensitivity to climatic factors at timberline.
Materials and methods
Study area
Study sites were located on the eastern slope of the Changbai Mountains(Fig.1,41.52°-42.47°N,127.15°-128.92°E), Jilin Province, northeastern China. This mountain range peaks at 2691 m a sl,making it the highest mountain in northeastern China,and it is a dormant volcano containing a vegetation community which has become established since the last eruption,approximately 1000 years ago(Liu et al.1993).Due to the height of the Changbai Mountains,an increase in global temperatures could have a significant impact on sustainability of the alpine ecosystem on this mountain(Wang et al.2015).The climate in the study area,being typically temperate continental,is affected by the East Asian monsoon,and it is characterized by long,cold and windy winters,and warm wet summers.Tianchi meteorological station is the nearest observatory,situated adjacent to the peak(Figs.1 and 5,42.03°N,128.07°E,2623.0 m a s l).Mean annual temperature recorded from 1959 to 2009 is-7.04°C;the coldest and warmest months are January and July,with mean temperatures of-22.89°C and 8.67°C,respectively.Mean annual precipitation is 1361 mm,of which about 62%occurs between June and August(Fig.5).
Climatic variations along elevation gradients on the Changbai Mountains are strong and clear vertical vegetation zonation is evident(consisting of mixed forest,conifer forest,birch forest and tundra),especially on the northern slope,which supports the major forest types in northeast China(Liu et al.1998;Yu et al.2011).However,vegetation on the eastern slope has weak divergence with elevation,and it is uniformly dominated by pure L.olgensis forest from the mountain foot to timberline(2000 m a s l).The difference in vegetation patterns between the north and eastern slopes is generally believed to be related to themajor volcanic eruptions(Dai et al.2008).Although the mechanisms for the development of the L.olgensis forest are not yet clear,it is acknowledged that the timberline of this forest was historically higher and that the current timberline is due to volcanic activities(Liu et al.1993).Since this area has had little anthropogenic disturbance,it is an ideal area to investigate tree growth-climate interactions(Lin et al.2013).
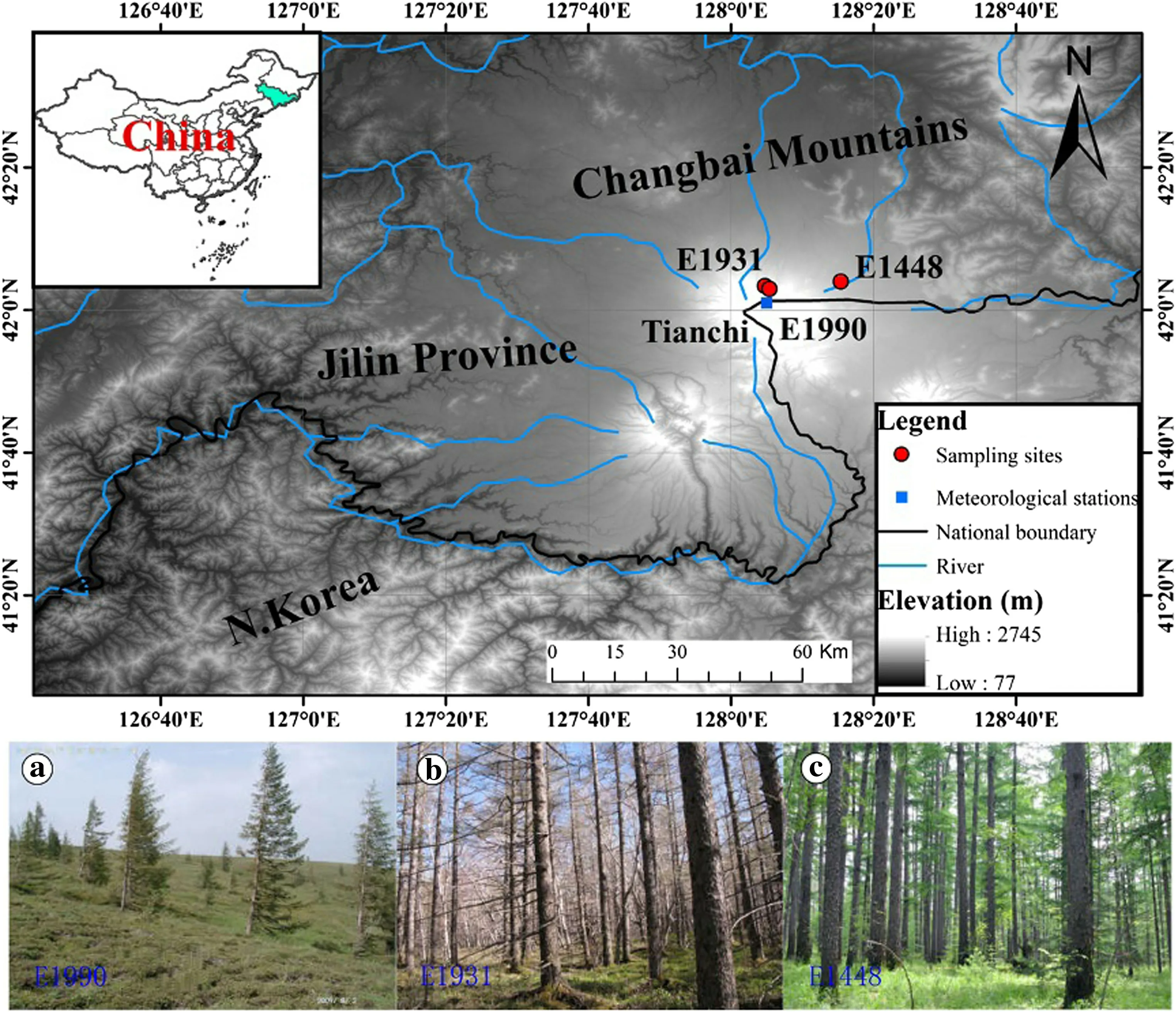
Fig.1 Map showing positions of sampling locations(red solid circle)and the meteorological stations(blue square)in the Changbai Mountains of Jilin Province,northeastern China.Photographs of forests along the altitudinal gradient on the eastern slope of the Changbai Mountains at the a upper elevation above the timberline(site code E1900),b upper elevation below the timberline(site code E1931)and c midelevation(site code E1448),respectively
Tree-ring sampling and chronology development
Tree increment cores of L.olgensis were collected at three sites along a vertical transect on the eastern slope of the Changbai Mountains, spanning an elevation of 1448-1990 m a s l.The three ranges were termed:(1)the upper elevation above the timberline(A:1990 m,hereafter referred to as‘‘E1990'');(2)the upper elevation below the timberline(B:1931 m,hereafter referred to as‘‘E1931'');and(3)the mid-elevation(C:1448 m,hereafter referred to as‘‘E1448'')(Fig.1 and Table 1).Core samples were collected in early May 2009 and August 2009 and 2010,all trees were visually assessed before sampling to ensure that only healthy trees were sampled.To minimize damage to the trees,only one core was extracted from each tree at breast height(1.3 m from the ground).In total,239 cores were extracted.Cores were transported to the laboratory and air dried prior to mounting and sanding,following standard dendrochronological techniques (Stokes and Smiley 1968).Cores with abnormal ring features were excluded from analysis,such as those missing rings or having indistinct boundaries which made cross-dating difficult.
Tree-ring widths were measured to 0.01 mm using the LINTABTm6.0 measuring system,the quality of measurement and cross-dating was checked using COFECHA(Holmes 1983).To remove any influence caused by nonclimatic factors,tree-ring width and biomass increment series were detrended and standardized using ARSTAN(Cook and Holmes 1996),using a 67%cubic smoothing spline function at a 50%cutofffrequency.To investigate the difference in tree-ring width chronology characteristics at our three different study elevations(Figs.2 and 3),the following statistical parameters were computed to compare the three chronologies:mean sensitivity(MS),standard deviation(SD),first-order autocorrelation(AC1),expressed population signal(EPS),the variance explained by the first principal component(PC#1),and signal-to-noise ratio(SNR)(Fritts 1976).The subsample signal strength(SSS)at a threshold value of 0.85 was used to identify the most reliable period of each chronology(Wigley et al.1984).Taking all these statistical methods into account, wefocused on residual chronologies for dendroclimatic analysis.
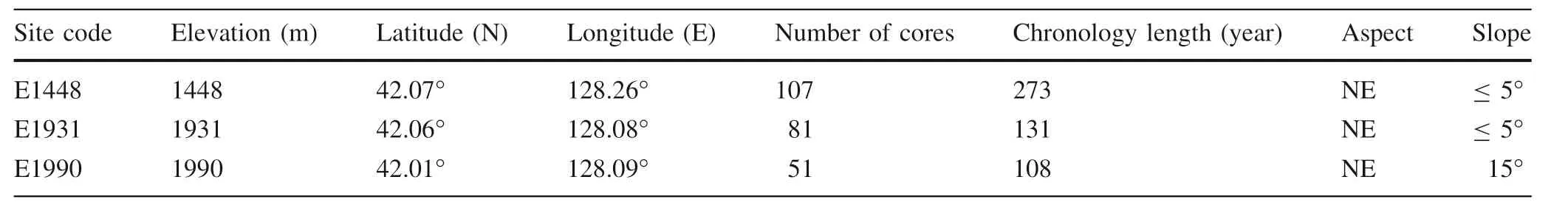
Table 1 Description of the three sampling sites in the eastern slope of the Changbai Mountains

Fig.2 Residual chronologies of tree-ring width along the elevation gradient.The red dashed line indicates the starting year of SSS(subsample signal strength)>0.85
Biomass increment
We used biomass increment to investigate responses to climate change because it is considered to be a more meaningful indicator of tree growth than tree-ring width(Fang et al.2015;Lin et al.2013).Stem biomass was estimated by converting stem volume to dry weight based on basic wood density(Eq.3),which is 0.505 g cm-3.Stem volume was estimated using a two-way volume equation(Ministry of Forestry in 1978)in the form of V=aDbHc(Eq.1).The volume equation was transformed to a one-way model by substituting item H with a DBHtree height model(Eq.2)which was established by the local forestry department.
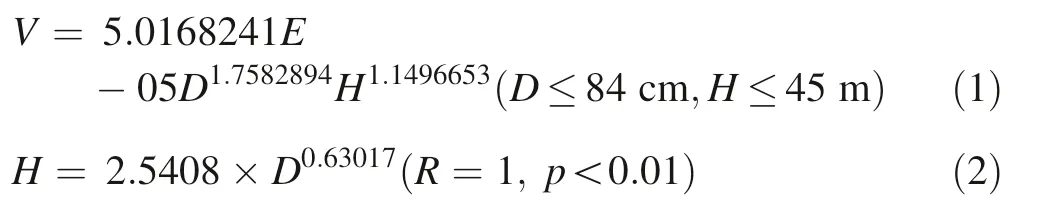

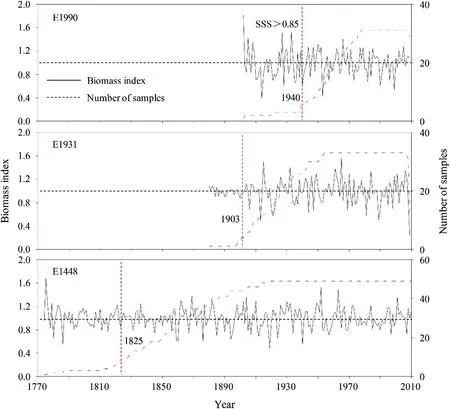
Fig.3 Residual chronologies of biomass along the elevation gradient.The red dashed line indicates the starting year of SSS(subsample signal strength)>0.85
where,H is tree height;D is caliper diameter at breast height;and V is stem volume(m3).where Wn(kg)is annual stem biomass increment at year n,Bnis individual stem biomass at year n,and Bn-1is individual stem biomass at year n-1.The annual stem biomass increment sequence of each tree was calculated using Eq.(4).
Cross-dating passed the inspection of tree-ring cores of 51,35 and 31,and ten cores were carefully chosen to analyze tree-ring width growth processes and annual biomass increments across the different elevations,respectively.These cores,with the pith,showed longer time spans than other samples.In addition,in order to examine the growth process, tree-ring widths and annual biomass increments were aligned in physiological age,a method that differed from traditional approaches for climate response analysis.Tree-ring widths and annual biomass increments were outlined based on accumulation and average values at the three sampling sites,respectively(Fig.4).
Climate data
Climate data,including monthly mean temperature(Tm),mean maximum temperature(Tmax),mean minimum temperature(Tmin)and precipitation(Pm)were obtained from Tianchi meteorological station(Figs.1 and 5),spanning the period 1959-2009.Distances from the station to the three sampling sites were 2.91 km for E1990,3.19 km for E1931,and 16.27 km for E1448.Climate data were provided by the China Meteorological Data Sharing Service System(http://cdc.cma.gov.cn/).To quantify drought conditions at the three sites,we used the Palmer Drought Severity Index(PDSI).This index is a direct metric of soil moisture conditions,and it uses surface air temperature,precipitation and evapotranspiration to indicate drought conditions.This index is believed to be a better indicator of drought conditions than solely relying on precipitation records(Dai et al.2004).The global PDSI dataset,updated by Dai et al.(2004),has a global resolution of 2.5°×2.5°spanning 1959-2012 and the geo-position of the nearest grid to our study area was 128°-128.5°E/42°-42.5°N.PDSI data for our study sites were downloaded from the KNMI climate explorer(http://climexp.knmi/nl).
Statistical analyses

Fig.4 The radial and stem biomass growing process of L.olgensis along the elevation gradient.Annual increment data and temperature were smoothed with 11-year running average
Response function analyses between tree-ring widths,biomass chronologies,and climatic factors were undertaken using DendroClim2002 software (Biondia and Waikulb 2004).A total of 1000 bootstrapped samples were used to compute response coefficients and to test their significance at the 0.05 level.Considering that tree growth could be influenced by climatic conditions in the previous and current year (Fritts 1976), we applied correlation analysis to a 17-month interval from May of the previous growth year to September of the current growth year from 1959 to 2009.We used intercorrelation among different chronologies to evaluate their degree of similarity and to detect inter-site differences. As the wavelet coherence method indicates local correspondence at different timefrequency domains,we used this method to evaluate similarities among chronologies(Grinsted et al.2004).Spatial correlation between tree-ring width,biomass chronologies,and the 0.5°×0.5°gridded meteorological datasets were generated using the KNMI climate explorer(http://www.climexp.knmi/nl),these maps enabled evaluation of the representativeness of chronologies at regional scales.
Results
Chronological characteristics
Descriptive statistics sharing a common interval analysis are listed in Table 2.Mean tree age at the three sampling sites ranged from 108 to 237 years and mean ring-width,which varied from 0.954 to 0.997 mm, significantly decreased with increasing elevation.Mean sensitivity(MS)ranged from 0.19 to 0.29 and increased with elevation,reflecting the high-frequency signals in the chronologies.MS also indicated that individual trees grew synchronously and that inter-annual variation in tree-ring widths was consistent at high elevation.Standard deviations and firstorder autocorrelations(AC)ranged from 0.18 to 0.24 and-0.06 to 0.10,respectively.The subsample signal strength(SSS)of chronologies at all sample sites exceeded 0.85.The chronologies for the three study areas were reliably dated to:AD 1938(E1448),AD 1903(E1931)and AD 1821(E1990).Among the three sites,trees below the timberline had the largest values of correlation(R)between trees,signal-to-noise ratio(SNR),expressed population signals (EPS), and variation in the first eigenvector(PC1%). Based on this analysis, we found that the chronologies for L.olgensis were rich in climatic information,thus making this species suitable for dendroclimatological investigations.
Growing processes and intercorrelations among chronologies
Comparison of radial growth showed that they were highly consistent across our three study elevations.The growth process of tree-ring widths was consistent with the trend of biomass increment(Fig.4).In general,L.olgensis rapidly grew during the juvenile period(0-40 years).This was followed by a steady decrease in growth rate from the halfmature to mature period(41-140 years),possibly caused by competition among trees. After 140 years, growth appeared to be robust as trees entered the over-mature period,a period which is considered the result of selfthinning and climate warming.The rapid growth of the over-mature stand illustrates that this old-growth forest was still vigorously sequestering carbon.
Intercorrelations,the similarity between tree-ring width and biomass chronologies at different time-frequency domains, are shown in Fig.6. For tree-ring widthchronologies,significantly positive correlations between E1990 and E1931 were found for periods of less than 20 years throughout the span from 1900 to 2009.However,E1448 was only significantly correlated with E1931 and E1990 in a few short intervals.For biomass chronologies,significant positive correlations between E1990 and E1931 extended from high-to low-frequency domains.Given that E1990 and E1931 showed a high degree of similarity among their chronologies,the chronology of E1448 was a remarkable discrepancy.

Table 2 Tree-ring width chronologies for L.olgensis along an elevational gradient in eastern slope of the Changbai mountains
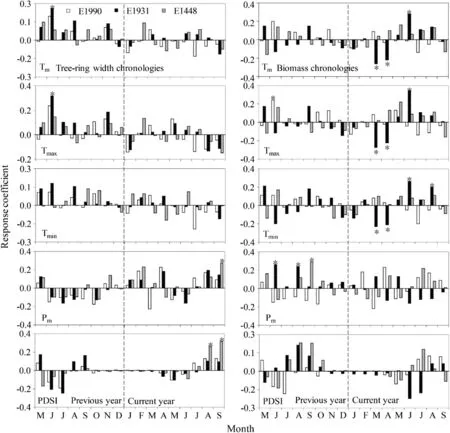
Fig.6 Response function analysis of tree-ring width(a)and biomass chronologies(b)for L.olgensis in relation to monthly climate factors at different elevations;*p <0.05
Climate-growth relationships along the elevation gradient
The response of tree-ring width chronologies to climatic changes are shown in Fig.6a.E1448 had a strong positive correlation with precipitation in September of the current year.E1448 was also significantly positively correlated with PDSI in August and September of the current year.Tree-ring width chronology responses at high elevations considerably differed from those at mid elevations.E1931 was less affected by summer hydroclimate conditions and it was only significantly and positively correlated with mean and maximum temperature in June of the previous year.In contrast,correlation with temperature and precipitation at E1990 showed less significance.In summary,tree-ring width chronologies of L.olgensis had a greater positive response to temperature at E1931 than at E1990.
The responses of biomass chronologies to climatic factors differed from the responses of tree-ring width chronologies(Fig.6b).The biomass chronologies at E1448 were only positively correlated with precipitation in September of the previous year.In comparison,biomass chronologies at E1931 had a more complex relationship with climatic factors;they were negatively correlated with air temperatures in March and April of the current year but positively correlated with air temperature in June of the current year.Biomass chronology was significantly and positively correlated with minimum temperature in August of the current year.Biomass chronologies at E1990 were also significantly positively correlated with maximum temperature in June of the previous year.
Large-scale drivers in the Changbai Mountains
To investigate the common growth forces from 1957 to 2009 in more detail,spatial correlations between tree-ring width,biomass chronologies,and the 0.5°×0.5°gridded temperature data were analyzed. Spatial correlations between tree-ring width chronologies at E1931 and the gridded surface temperature data revealed that temperature changes in northeast China were recorded,with the highest correlations occurring in the Changbai Mountains(Fig.7a).Spatial correlations derived from our biomass chronologies at E1931 and the CRU gridded dataset generally show a similar pattern(Fig.7b),and spatial correlation results confirmed that our chronologies recorded broad-scale regional climatic variations across most of northeastern China.Results from E1931 showed potential to reconstruct summer climates using biomass chronologies,but results from E1990 to E1448 did not.
Discussion
Chronology strength
Our analysis provided an effective tool to understand the relationship between radial growth and climatic factors.Our results were consistent with previous investigations for this species(Chen et al.2011;Shen et al.2015)and other species(Yu et al.2005a,2007)in the same region.Our results suggest that L.olgensis is among those tree species that are very sensitive to climate change,and that its trees contains abundant climate signals.These findings accord with those of Chen et al.(2011).Furthermore,the sensitivity to climatic changes of the chronologies at high elevations was greater than that at mid-elevations.Shen et al.(2015)reported that trees near the limit of the species distribution,rather than those within the inner forest,were suitable for climate-growth studies.
Our results showed a decreasing trend of mean tree-ring width with elevation,a finding frequently reported for mountain areas(Peng et al.2008;King et al.2013).This was probably due to the typically lower temperatures and shorter growing seasons at higher elevations.Lower temperatures and shorter growing seasons at high elevations may result in trees showing low metabolic rates as they adapt to the environment(Liang et al.2010).Although the strongest climatic control on tree growth is expected in forests located at the limit of their species'distribution(Fritts 1976),our highest-elevation sampling site showed relatively poor agreement between trees. This finding supports the results reported by Kharal et al.(2017)from studies in central Nepal.Above timberline,tree-ring formation is limited by various factors,such as low temperature,strong wind exposure,shallow soil,low nutrient availability,and short growing season(Liang et al.2010).Additionally,in the ecotone at the tree line,the greater wind speeds at the perimeter of tree stands can cause structural damage to trees,especially in the tree crown(flag tree),and this affects radial growth(McIntire et al.2016).
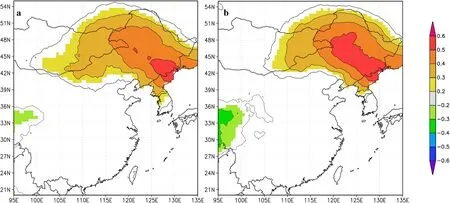
Fig.7 Spatial correlations between tree-ring width(a)and biomass chronologies(b)and the grid data of climate factors from CRU TS 4.01 land dataset over the period AD 1959-2009
Climate-growth relationship along the elevation gradient
Response function analysis between climatic factors and tree-ring width supports the hypothesis that climate-growth relationships vary along elevation gradients (Fig.6), a pattern reported by Chen et al.(2011)in the Changbai Mountains.Similar findings have also been reported for the same tree species in northeast China(Shen et al.2015;Zhuang et al.2017)and for other tree species at high elevations in the Northern Hemisphere(Andreassen et al.2006;Peng et al.2008;King et al.2013).Correlation analysis showed that radial growth at E1990 and E1931 exhibited similar responses to climate,while radial growth at E1448 responded differently.This finding was validated by the climate response analysis,in which chronologies showed positive responses to summer temperatures at E1990 and E1931,and to precipitation at E1448(Fig.8).
Temperature, particularly in June, is a pronounced growth-limiting factor for L.olgensis at the timberline on the eastern slope of the Changbai Mountains.Our results support those reported for high-elevation Korean pine and subalpine L. olgensis chronologies across the northern slope of the Changbai Mountains(Chen et al.2011;Shen et al.2015;Zhuang et al.2017).Asad et al.(2016)also found temperature,rather than precipitation,to be the fundamental constraining climatic factor for tree growth towards the upper tree line in the Karakoram.Tree growth at cold,moist sites at high latitudes also shows strong correlation with temperature variability over large areas of the Northern Hemisphere(Dulamsuren et al.2011;Chen et al.2012;Jiang et al.2016;Zhang et al.2016).In northeastern China,the short growing season near timberline spanned only July and August;higher summer temperatures promoted earlier snow-melting and more rapid soil warming,thereby lengthening the snow-free growing season.In addition,higher temperatures in June would increase leaf area and the photosynthetic rate of trees.In contrast,low summer temperatures at timberline might limit cambial activity in early spring and cause damage to needles and shoots from spring frost,or even damage cambial tissue.
Mid-elevation sites were warmer than high-elevation sites;the positive influence of temperature on tree growth disappeared,and temperature exerted a negative effect on tree growth at the mid-elevation sites.The radial growth of L.olgensis at mid-elevation was strongly restricted by precipitation,especially towards the end of the growing season(September in particular),indicating that trees at lower elevations suffered more from drought stress than did trees at higher elevations.Previous investigations on lower forest borders in northeast China and from semi-arid regions showed that precipitation influences tree growth(Yu et al.2007,2013;Cai and Liu 2013).In contrast,water deficit rarely limits alpine or high-elevation forest growth.In the Changbai Mountains,L.olgensis is a photophilous and drought-tolerant tree species,but its growth rate is closely related to soil water and nutrient conditions.Precipitation during the end of the growing season exerts an important influence on tree growth on the lower forest border(Shen et al.2015).Despite relatively high annual rainfall at our study area and about 62%of precipitation occurring from June to August in the growing season,precipitation levels during September were low,and this limits tree growth and assimilation processes.In general,higher summer temperatures,coupled with low levels of precipitation,accelerate transpiration and soil evaporation which results in strong moisture stress on tree growth,thereby inhibiting tree-ring growth.In contrast,adequate precipitation in September can extend the growing season,which enhances the accumulation of carbohydrates,soil moisture conservation,and the formation of wide treerings.The significant positive correlations of tree growth with PDSI at the end of the growing season further confirm the important role moisture plays in regulating the growth of L.olgensis.Tree growth at the lower-elevation sites was mainly limited by moisture availability.
The sensitivity of biomass chronology
Our results show that biomass chronologies were capable of reflecting climate regime,findings that support previous reports for the Changbai Mountains(Fang et al.2015).Lin et al.(2013)reported that the relationships between annual biomass increment,radial growth and biomass chronologies can be effectively applied in dendroclimatology research.In comparison with tree-ring width chronologies,biomass chronologies showed stronger responses to climatic factors at all of our sampling sites.This suggests that biomass chronologies are more suitable for capturing the effects of climate signals.
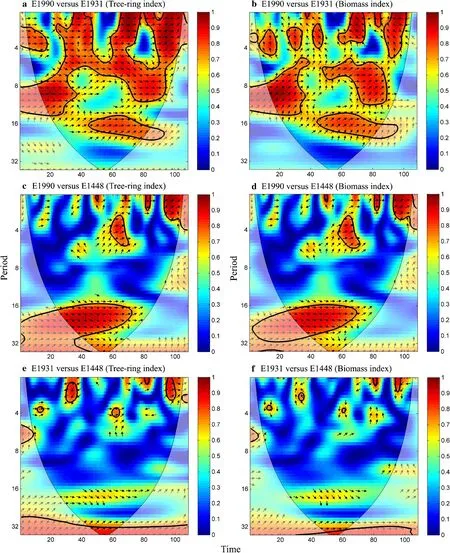
Fig.8 Wavelet coherence among the tree-ring width and biomass chronologies from 1902 to 2009.Arrows indicate the phase of the coherence,where right arrow is in phase and left arrow is antiphase.The 95%confidence level against red noise is shown as a black contour
Our results show that biomass chronologies and early spring air temperatures(March and April)of the current year were significantly negatively correlated at high elevation(Fig.6).These findings accord with those of Chen et al.(2011)and Shen et al.(2015)reported for L.olgensis at the same area.Shao and Wu(1997)also showed that warm temperatures in March and April were unfavorable for tree growth.At high elevations,winters are generally cold and long,and March mean temperatures are normally below-20°C(Fig.5).Therefore,a warm March would not result in an earlier activation of the cambium,but instead would consume the photosynthetic reserves of the previous year.In contrast,positive correlations between tree growth and temperature during the previous winter were reported for other tree species(Dang et al.2013).These differences may result from inherent diversity of growth patterns among tree species and specific climatic conditions.
Decreased sensitivity of tree growth above timberline
The complex relationship between tree-ring width and climatic factors has been widely reported,particularly in extreme environments in high mountains and the subarctic zone.The climate at the forest margin being the main limiting factor of tree growth(Korner and Paulsen 2004;Toledo et al.2011).If trees growing at their elevation limit are most sensitive to climate change,tree growth-temperature relationships should be strongest towards the tree line,a conclusion which has been widely confirmed(Yu et al.2005a;Liang et al.2010).
Results from trees located near the tree line in our study,however,differ from the general assumption that trees growing at or near the tree line are more sensitive to temperature than trees growing at lower elevations.This result accords with the findings of Cai and Liu(2013)and Kharal et al.(2017)who recorded a similar lack of sensitivity at the highest elevations in the Lu¨liang Mountains of northern China,and in central Nepal,respectively.The weak correlation is possibly a result of competition,or the impact of natural disturbance, which accentuates the deviation at the tree line,thereby reducing climate sensitivity(Norton 1985).In addition,exposure to strong winds,shallow soil and low nutrient availability might also contribute to decreased tree growth at high elevations which will lead to reduced dendroclimatic sensitivity(Liang et al.2010).Wind is considered to be a frequent natural disturbance which affects the forest structure and its dynamics,especially for coniferous forests at higher elevations(Mcintire et al.2016),as shown by the flag tree in Fig.1a.Larch forests located below timberline at our study area experienced no climatic disturbance which would capture the true relationship between tree-ring width and climate factors,and the cores sampled here may provide more climatic information. Therefore, the optimal site for examining tree-growth trends in response to climate variation might be located below timberline(Norton 1985;Cullen et al.2001).
Based on these analyses,we conclude that there was an obvious elevation effect on radial growth patterns and climate-growth relationships in our study area.The relatively cold and moist conditions on the special topography of the eastern slope of the Changbai Mountains,together with the wide elevation range of L. olgensis forests,probably resulted in elevation-dependent climate-growth relationships.However,the explanatory power of the climate-growth relationships we calculated might have been limited by the available weather station data,thus further studies are needed to verify our results in this study area.
Conclusions
Our results suggest that elevation affects tree growth patterns in this mountainous region.Tree growth at high altitude is strongly limited by June temperatures of the previous year and,with decreasing altitude,the importance of temperature decreased;Mid-elevation tree radial growth was mainly controlled by precipitation towards the end of the growing season of the current year.In comparison with tree-ring chronologies, biomass chronologies showed stronger responses to climatic factors at all sites.Spatial correlation with gridded climate data revealed chronologies containing a strong regional temperature signal for northeast China.As trees growing below the timberline appeared to be more sensitive to climatic warming,optimal sites for examining growth trends as a function of climate variation are therefore considered to be just below the timberline.These results provide new information on growth responses of L.olgensis to climate change on the eastern slope of the Changbai Mountains that might be used in future climate reconstructions using this tree species in humid and semi humid regions.
AcknowledgementsAuthors express their thanks to Sher Shah,Qianqian Xu,Guochun Zhang,Junlong Yang,and Wenhui Liu for their help with field sampling and core measurement.Authors also thank the reviewers for their diligence in providing useful comments that improved this work.
Author's contributionsJian Yu and Qijing Liu conceived the idea;Jian Yu and Qijing Liu sampled the tree cores and measured the treering widths;Jian Yu conducted the analysis and wrote the manuscript.
杂志排行

Journal of Forestry Research的其它文章
- Protective and defensive roles of non-glandular trichomes against multiple stresses: structure-function coordination
- Assessment of early survival and growth of planted Scots pine(Pinus sylvestris)seedlings under extreme continental climate conditions of northern Mongolia
- Influencing in vitro clonal propagation of Chonemorpha fragrans(moon)Alston by culture media strength,plant growth regulators,carbon source and photo periodic incubation
- Variation analysis of growth traits of four poplar clones under different water and fertilizer management
- Nodule study in Albizia chinensis in relation to nitrogen metabolism,morphology and biomass
- Comparative transcriptome analyses reveal candidate genes regulating wood quality in Japanese larch(Larix kaempferi)