The mechanism underlying overwintering death in poplar:the cumulative effect of effective freeze-thaw damage
2020-01-18ChengchaoYangJiandongPengXiaoyuLiDejunLiangZhiyanYangYanZhang
Chengchao Yang·Jiandong Peng·Xiaoyu Li·Dejun Liang·Zhiyan Yang·Yan Zhang
Abstract We analyzed the relationships linking overwintering death and frost cracking to temperature and sunlight as well as the effects of low temperatures and freeze-thaw cycles on bud-burst rates,relative electrical conductivity,and phloem and cambial ultrastructures of poplar.Overwintering death rates of poplar were not correlated with negative accumulated temperature or winter minimum temperature.Freeze-thaw cycles caused more bud damage than constant exposure to low temperatures.Resistance to freeze-thaw cycles differed among clones,and the budburst rate decreased with increasing exposure to freezethaw cycles.Cold-resistant clones had the lowest relative electrical conductivity.Chloroplasts exhibited the fastest and the most obvious reaction to freeze-thaw damage,whereas a single freeze-thaw cycle caused little damage to cambium ultrastructure.Several such cycles resulted in damage to plasma membranes,severe damage to organelles,dehydration of cells and cell death.We conclude that overwintering death of poplar is mainly attributed to the accumulation of effective freeze-thaw damage beyond the limits of freeze-thaw resistance.
Keywords Effective freeze-thaw·Freeze-thaw resistance·Mechanism·Overwintering death·Populus·Ultrastructure
Introduction
Poplar,of the genus Populus in the Salicaceae family,is one of the most important landscaping and timber tree species in the plains.In northern China,the fast-growing Aigeiros poplar has gradually replaced native poplar and has become the predominant species.However,in Liaoning,Hebei,Inner Mongolia and other northern provinces,fast-growing poplars often suffer from winter injury,such as that caused by freezing.Liaoning Province,a typical region where winter injury to poplar occurs,is located in the southern part of the Northeast China Plain,in the transition zone between the warm and temperate zones,where the climate is variable.Over the past decade,large areas of dieback and overwintering mortality have been recorded many times in Liaoning Province.
Winter injury in trees involves visible damage or death of organs,tissues or whole trees and is directly caused by stressful winter conditions, including freezing injury,photo-oxidation,and other factors when the daytime minimum temperature is below 0°C(Tao and Jin 2005).Overwintering death is an extreme outcome of overwintering injury.Studies on winter injury have focused on red spruce (Picea rubens) (Strimbeck et al. 1991, 1993;Strimbeck and Dehayes 2000),apple(Malus pumila)and other species(Larcher 1981;Pramsohler et al.2012).
Freezing injury results from damage caused by temperatures below freezing;for temperate trees,frost resistance is key to surviving the winter(Ko et al.2011).The known mechanisms of frost damage in trees involve irreversible systemic changes(Asada and Kanematsu 1976;Steponkus 1984;Hellergren et al.1987;Webb et al.1994),the presence of intercellular and intracellular ice(Ristic and Ashworth 1993b)and structural damage to cell membranes caused by exposure to extremely low temperatures below freezing(Sakai and Larcher 1987)or to low temperatures for long periods of time(Lyons 2003),ultimately leading to cell dehydration and death.Tao and Jin(2005)studied the frost resistance of 51 woody plants and found that the frost resistance of trees exhibits seasonal variation and that there are significant differences in frost resistance in different organs and tissues.Species with stronger frost resistance,such as red pine(Pinus koraiensis),tolerated temperatures of-196°C,whereas aspen(P.tremuloides)branches and buds tolerated temperatures of -60°C.However,in the natural environment,winter death can occur even when low temperatures are higher than the frost-resistance threshold. Other factors also influence overwintering death.Freezing damage in forest trees is not restricted to extremely low-temperature conditions(Cox and Zhu 2003);freeze-thaw cycles are associated with forest dieback in northern hardwoods (Auclair et al.1996,1997).Groß et al.(1991)reported that freeze-thaw cycles increase winter injury in Norway spruce(Picea abies).It is currently unclear whether overwintering death in poplar is caused by exposure to extremely low temperatures or freeze-thaw cycles.
In addition to temperature,sunlight has an effect on overwintering injury to trees.Photoperiod alone is thought to determine the end of the growing season in poplar(Howe et al.1996;Bo¨hlenius et al.2006;Fracheboud et al.2009).However,even in trees thought to be photoperiod controlled, temperature can modify the timing of key phenological events(Mo¨lmann et al.2005;Fracheboud et al.2009;Tanino et al.2010).Strimbeck et al.(1993)reported that low temperature and strong sunlight can cause needle decoloration,chloroplast decomposition,and can inhibit photosynthesis(Strimbeck et al.1993)in red spruce foilage.However,the effect of light on overwintering death in poplar remains to be studied.
An important aspect of the mechanism underlying winter injury is the initial site of cell injury.There is a general consensus that the initial primary site of freezing in the cell is the plasma membrane.Plasma membrane damage is caused by osmotic stress and cell dehydration during freezing,and the plasma membrane is the primary site of the cellular response to freeze-thaw stress (Steponkus 1984).Studies of the relationship between organelles and winter injury have mainly focused on structural changes to the chloroplast during cold acclimation(Guy et al.1992;Risticand Ashworth 1993a).After cold acclimation,the number and size of starch grains in chloroplasts increase,chloroplasts enlarge, vacuole size is reduced, and cytoplasmic volume increases(Strand et al.1999;Nagao et al.2008).As structural changes in the cell are associated with freezing resistance,it is important to study the ultrastructural changes in the cells of poplar suffering winter damage.
In this study,we analyzed the relationship linking poplar winter mortality and trunk frost cracking to temperature and sunlight over a 5-year period in the field.In addition,using laboratory experiments,we studied the effects of low temperature and freeze-thaw cycles on bud-burst rates,relative electrical conductivity and cellular ultrastructure of poplar.Extremely low temperatures are considered the root cause of freezing injury(Tao and Jin 2005).Our aim was to determine whether the lowest temperatures or effective freeze-thaw cycles caused poplar death of Linghai,Liaoning Province in the winter.We assumed the following:(1)overwintering death in poplar is associated with frost resistance;(2)overwintering death in poplar is associated with freeze-thaw cycles;(3)freeze-thaw cycles have significant effects on cellular ultrastructure.
Materials and methods
Field trials
Description of study sites
The field trials were performed at a monitoring station in Linghai, Liaoning Province, China (N41°12′8.65′′,E121°24′1.21′′,elevation 16.85 m).Average annual temperature ranges from 8 to 8.7°C,≥10°C accumulated temperature is 3400-3600°C, and annual rainfall is 550-620 mm,with 160 to 180 frost-free days.
Test design
An experimental plantation of poplar was established using 1-year roots without stems of P.×canadensis‘Guariento',P.×euramericana‘Zhongliao1',P.×deltoides‘Jing2'and P.×euramericana ‘N3016' in April 2007. The plantation was planted with 6 m between rows and 2 m between adjacent individuals in each row.Three blocks served as biological replicates in a randomized block design.Each block contained 110 plantlets of each clone.
A tree was marked as the injured tree and was no longer used as the investigational object in the next year if it was suffered from wintering injury. In order to meet the requirement of sample number,some nearby healthy trees of these four clones equal in age to those in our plantation were selected as supplementary to survey winter injury.Before winter,in totals 100 healthy trees with no white paint for each clone were randomly selected and marked in the experimental plantation and supplementary plantation.The overwintering death rates(DRs)of these trees were investigated in June of the second year from 2008 to 2012.Death of an individual was defined as death of at least 50%of the stems.
In order to study winter injury of 1-year-old plantlets,1-year-old plantlets of the four clones were propagated from hardwood cuttings.The breeding number of plantlets for each clone was 3000 per year from 2008 to 2012.Each April from 2009 to 2013,the DRs of 1-year-old seedlings were investigated.In total,100 seedlings of each species were randomly sampled.The DR was calculated on the basis of the seedlings.
In order to illuminate the mechanism of the trunk frostcrack,whitewash whose main component was CaCO3and Ca(OH)2was painted on tree trunks from ground level up 1 m on 50 trees per block each October from 2008 to 2012.Trees with no white paint served as a control group.Each June from 2009 to 2013,the trunk frost-crack rates of these four clones were investigated.
The air temperature and the temperature of the cambium and bark of shaded and sunlight-exposed,painted and unpainted P.×deltoides‘Jing2'were monitored using an NTC(thermistor sensor)ZDR-20 temperature recorder(-made in China). Recording was conducted from 1 November 2008,to 31 March 2013,with measurements taken every 30 min.The diurnal temperature range(DTR)represents the difference between the maximum and minimum temperatures.We recorded the DTR of air,shaded bark,and sunlight-exposed bark;the monthly average air temperature;the annual negative accumulated temperature,and average winter temperature. The lowest monthly temperature in winter in each year was recorded.Pearson correlation analysis was used to describe the relationships between the annual negative accumulated temperature,winter minimum temperature,winter average temperature and stem frost crack rate,large-tree DR,and seedling DR.We used IBM SPSS Statistics22 for this and all other statistical analyses.Our alpha level for all analyses was 0.05 unless otherwise noted.
The diurnal variation curves for air temperature,cambium temperature,and bark temperature on the shaded and sunlight-exposed sides of painted and unpainted poplar were plotted based on the data from days that had adequate sunlight and low temperature.
Laboratory testing
The genotypes
One-year-old branches of cold-resistant and non-cold-resistant clones of 11 cultivars were used as test materials in late March before poplar bud burst, including P.simonii×P.nigra,P.simonii Carr.,P.×euramericana‘Zhongliao1',P.deltoids×(P.nigra var.thevestina×P.nigra),P.×canadensis‘Guariento',P.×deltoides cv.‘Liaoyu3', P.×euramericana ‘97', P.×euramericana‘L35', P.×xiaozuanica ‘Poplar', P.×canadensis‘Sacran-79'and P.×euramericana‘N3016'.
Treatment of test materials
Freezing treatment For each clone,the stems of 33 similarsized 1-year-old seedlings were sampled and divided into 11 groups.Each clone had 3 shoots in a group,or 3 repetitions.Each shoot had over 30 buds.The materials were placed in a cryogenic refrigerator and exposed to temperatures of 0 to-50°C over a 5°C gradient;the cooling rate was 2°C h-1,with 0°C used for the control(CK).After exposure to the required temperature for 24 h,some materials were subjected to growth and recovery treatment,whereas the remaining branches were subjected to additional cold treatment.
Freeze-thaw treatment For each clone,the stems of 15 1-year-old, similar-sized seedlings were sampled along with the tops of 30-50-cm seedlings.The materials were divided into five groups.Each clone had 3 stems and tops of seedlings in a group,for 3 repetitions.Each stem and top had over 30 buds.For each cycle,the materials were exposed to a temperature of-40°C for 24 h,followed by 20°C for 24 h.A group of materials was subjected to growth and recovery after each cycle,and the remaining materials were exposed to additional freeze-thaw cycles for a total of five freeze-thaw cycles.A room-temperature treatment was used for the CK.
Growth recovery
The materials were cut into 15-20-cm-long segments and placed into plant growth chambers for subsequent hydroponic treatment.The water was changed every 3 days.The materials were grown at 25±2°C under a 14 h/10 h light/dark cycle.The bud-burst rate of each clone was calculated after 28 days.
With the bud-burst rates as the dependent variable,the number of freeze-thaw cycles and the clones as the fixed factors,univariate,3-replicates,and two-way analysis of variance(ANOVO)was performed.
Two-way ANOVA, one-way ANOVA, and Turkey multiple comparisons were performed for each clone.The purpose of this study was to examine the small differences in bud-burst rates among freeze-thaw cycles.
Measurement of relative electrical conductivity after freeze-thaw treatment
According to the growth recovery of materials subjected to the two treatment methods,the branches of plants subjected to freeze-thaw treatment four times were selected based on the bud-burst rates to measure relative electrical conductivity(REC)(Wang et al.2003).Correlation between REC and bud-burst rates after four freeze-thaw cycles was quantified.
Ultrastructure of cambium and phloem after freeze-thaw treatment
Trees die when the cambium dies;therefore,it is necessary to study the ultrastructure of phloem and cambium cells before and after freezing and thawing.The top branches of 1-year-old P.×euramericana ‘Zhongliao1' (moderate cold tolerance)after freeze-thaw treatment were used as test materials.The cambium and the phloem were removed from the top 20 cm of each test material and cut into Sects.1 mm wide and 2 mm long at three time points:before treatment,after the first freeze-thaw treatment and after the fourth freeze-thaw treatment.Eight samples were taken per treatment,with two replicates.The materials were fixed in 2.5%glutaraldehyde in phosphate buffer(pH 7.2),followed by a wash in 0.1 mol phosphate buffer.The samples were then fixed in 1%osmic acid;washed in 0.1 mol phosphate buffer;dehydrated in 30%,50%,70%,80%,90%and 100%acetone;and embedded and polymerized in SPURR epoxy resin.Cambium localization was conducted using semi-thin sections.The samples were then sliced into ultrathin sections with a LEICAUC6i ultramicrotome,subjected to uranyl acetate and citric acid-lead double staining,and observed and photographed using a JEM-1230 transmission electron microscope.
Results
Field observations
Negative accumulated temperature,winter average temperature,trunk frost crack and overwintering death in poplar
Negative accumulated temperature was calculated as the sum of the daily average temperatures during a period in which the average daytime temperature was less than 0°C.The 5-month period of negative accumulated temperature at Linghai was from 1 November 1 to 31 March.Negative accumulated temperature is an important indicator that reflects the degree of exposure to cold temperatures;a low negative accumulated temperature indicates low temperatures in winter and a long period of low-temperature exposure.
The annual negative accumulated temperature was low in 2012-2013(Table 1).During this cold winter,poplar trees(DR=41.3%)and seedlings(DR=28.8%)died over a large geographic area.In 2011-2012,the annual negative accumulated temperature was the lowest of any winter in the 5-year period,but poplar death did not occur(DR=0).The annual negative accumulated temperature was at its highest in 2008-2009.Although this winter was warm,poplar trees(DR=54.5%)and seedlings(DR=37.2%)experienced massive death and trunk frost crack. The similar results(Table 1)existed in the winter average temperature too.Winter average temperature in 2008-2009 and 2010-2011 were high,but seedlings died a lot.
Annual negative accumulated temperature was not correlated with stem frost crack rate(r=0.440),large-tree DR(r=0.459),or seedling DR(r=0.575).Similarly,mean winter temperature was not correlated with stem frost crack rate(r=0.327),large-tree DR(r=0.348),or seedling D R(r=0.533).
Cold winters did not necessarily cause poplar death but warm winters led to massive overwintering deaths.There was no clear causal relationship between negative accumulated temperature and poplar overwintering death.This suggests that the poplar overwintering death recorded in recent years might not have been caused by exposure to low temperatures or long periods of cold temperatures.
Although the annual negative accumulated temperature of 2010-2011 was higher than during other study years,large trees did not die(DR=0)but among seedlings we recorded dieback and whole-seedling death(DR=32.6%).
Lowest temperatures and poplar death in the winter
The lowest temperature during our 5-year study period,-29.2°C (Table 1), was recorded during winter 2009-2010 when no poplar mortality was recorded.We documented no correlation between winter minimum temperature and stem frost crack rate(r=0.790),largetree DR(r=0.798),or seedling DR(r=0.464).Thus,the large-scale poplar winter mortality recorded during recent years was unrelated to winter minimum temperatures.
Freeze-thaw and trunk frost crack
More than 95%of poplar trunk frost cracking occurred in the southwest of trunk,where trunks were exposed to strong sunlight.The degree of injury was also closely related to the clone type.In June 2013,the trunk frost crack rates of P.×canadensis‘Guariento'and P.×deltoides‘Jing2'were both over 80%,whereas the trunk frost crackrate of P.×euramericana‘Zhongliao1'was 15%and there was no trunk frost crack in P.×euramericana‘N3016'.

Table 1 Overwintering injury of poplar and negative accumulated temperatures etc.from 2008 to 2012 in Linghai,China
From 9:00 to 16:00,the temperatures of sunlight-exposed cambium and bark were higher than those on the shaded side and that of the air(Fig.1a-d).Other time points yielded the opposite result,except that the air temperature was higher than that of both sunlight-exposed bark and shaded bark.In contrast to the temperatures at morning and at noon,the temperature of the cambium was higher than that of the bark from 14:00 to 16:00.This caused internal expansion and external contraction of the trunk(Fig.1c),possibly leading to separation of the bark from the xylem.The DTR of sunlight-exposed bark and cambium was nearly 35°C on 7 February 2009,indicating that the bark had undergone considerable freezing and thawing.Severe freezing and thawing caused serious damage to the trees(Fig.1e).In the environment of slight freeze-thaw and the same low night temperatures on the sunny side,no frost crack occurred in the shaded trunks,indicating that low temperature was not the main cause of trunk frost cracking.Rather,the cause was repeated severe freezethaw(Fig.1).Supporting this point,whitewashing prevented trunk frost cracking in P.×deltoides ‘Jing2'(Fig.1e).White paint reduced the temperature of sunlightexposed bark by reflecting sunlight and the heat radiation of soil,thereby reducing the DTR(Fig.1c,d;DTR was reduced by approximately 7°C by the paint)and preventing intense freeze-thaw damage.
The average temperature and the average DTR of sunexposed bark were both higher than the average air temperature(Table 2).In the winter,the average DTR of sunexposed bark reached 26.5°C and maximum DTR was 38.6°C.
Laboratory test results
The effects of different treatments on poplar
Through graded cooling,we determined that the bud-burst rates did not differ by clone at low temperatures.The budburst rates within individual clones did not vary by temperature,remaining >90%after 24 h at-50°C.
Bud-burst rates decreased significantly with increasing numbers of freeze-thaw cycles for all clones(P <0.01;Table 3).The bud-burst rates of the test materials were higher after the first freeze-thaw and did not vary among clones.The bud-burst rates decreased significantly after the second and third freeze-thaw cycles.
The resistance varied among clones to the number of freeze-thaw cycles,and the bud-burst rates of the clones after four freeze-thaw cycles differed significantly(P <0.01).The interaction between freeze-thaw cycles and clones on bud-burst rates was significant(P <0.01).
REC after freeze-thaw treatment
The 11 clones exhibited different degrees of damage after four freeze-thaw cycles.The RECwas significantly different among clones(Table 3)and was negatively correlated(r=-0.9101**,P <0.01)with the bud-burst rate.The bud-burst rate decreased with increasing exposure to freeze-thaw cycles,which seriously damaged the poplar branches.
Ultrastructure following freeze-thaw treatment
The first indication of winter injury was a change in the color of the phloem cells from yellowish white to brown,then black.
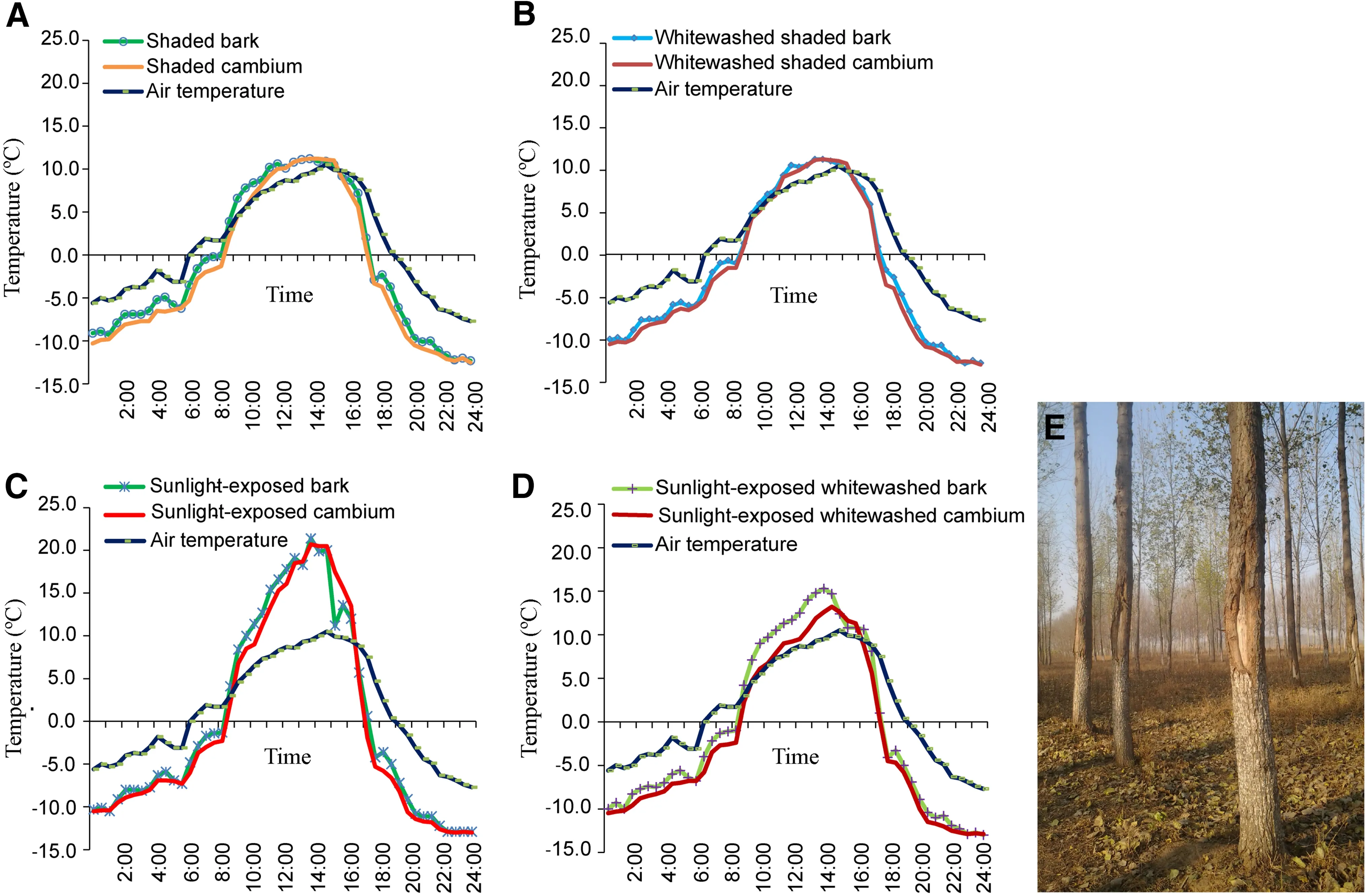
Fig.1 Daily variation in air temperature and the temperature of cambium and bark of both the shaded and sunlight-exposed side in P.×deltoides‘Jing2'that were painted white and not painted white on February 7,2009.c Shows that the temperatures of cambium and bark were not synchronized;the sunlight-exposed cambium temperature was higher than that of the sunlight-exposed bark from 14:00 to 16:00.The result was internal expansion and external contraction,which caused the bark to separate from the xylem.This process only slightly affected shaded bark.The DTR of sunlight-exposed bark was greater than that of shaded bark.c,d show that white paint could significantly reduce the temperature of sunlight-exposed bark and cambium during the day but did not affect the lowest temperature at night. a, b Show that white paint had little influence on the temperature of shaded bark and cambium.e The effect of white paint on P.×deltoides‘Jing2'.In the next year,the trunks painted white(up to 1 m above the ground)did not experience frost cracking,but on the southwest exposure,frost crack occurred from 1 m to 1.6 m above the ground,exposing the xylem.The southwest-facing trunks of P.×deltoides‘Jing2'(such as the two trees on the right and rear)that were not painted white all experienced frost cracking from the ground to 1.6 m;there was no frost crack on any other side.The effect of white paint on frost cracking was obvious
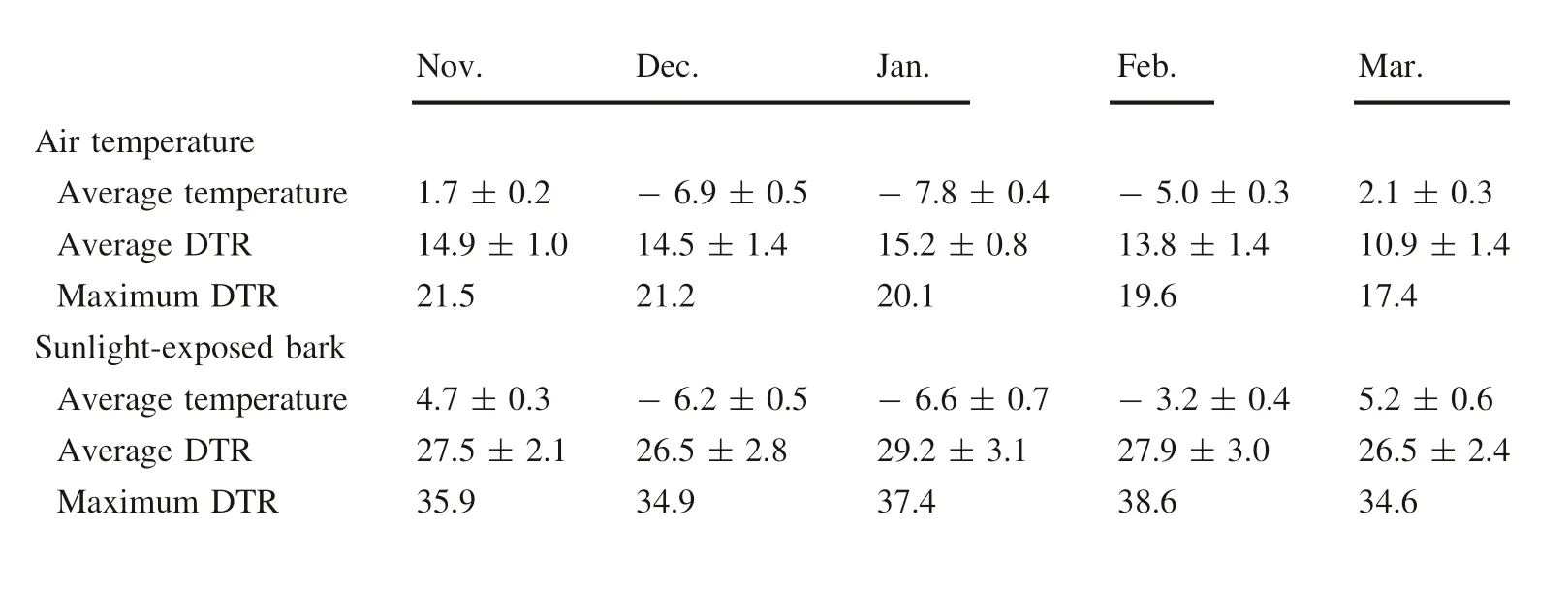
Table 2 Monthly variation in DTR in air temperature and temperature of sunlight-exposed bark of P.×deltoides‘Jing2'in the winter of 2008-2009
Before the freeze-thaw cycle,the cambium cell wall was thinner and the vacuole,nucleus and endoplasmic reticulum were visible(Fig.2a,b),whereas the chloroplasts were not.The mitochondrial cristae of the phloem parenchyma cells were clearly visible(Fig.2c),as was the rough endoplasmic reticulum(Fig.2c).The chloroplast has a complete double envelope and thylakoids arranged in parallel in the matrix(Fig.2d).

Table 3 Bud-burst rates after freeze-thaw and REC after four freeze-thaw cycles in poplar
After one round of freezing and thawing,the granum lamella of the parenchyma cell expanded and appeared dispersed. The chloroplast membrane was degraded(Fig.2g,h),and the lamellar structure disappeared.The mitochondrial membrane was degraded and hollowed(Fig.2e).After four rounds of freezing and thawing,the cell membrane(Fig.2i)was degraded.The nucleus was swollen,and organelle boundaries were not clear(Fig.2i).Severe dehydration(Fig.2j)caused irreversible plasmolysis,leading to cell death.After a cambium cell underwent freezing and thawing,the ultrastructure responded similarly,though there was no chloroplast.
The order of response to freezing and thawing exhibited by chloroplasts,mitochondria,and the endoplasmic reticulum are shown in Fig.2e,f.The chloroplast exhibited the fastest and most obvious response to freezing and thawing.
Conclusions
The overwintering death among poplar seedlings was inconsistent with that among large trees.The high DTR values of sun-exposed bark seriously affected the survival rate of poplar in winter.Repeated freeze-thaw cycles were the main cause of stem frost crack and tree death of Linghai,Liaoning Province in poplar.
The results of laboratory test indicated that freeze-thaw cycles caused more bud damage than constant exposure to low temperatures.Chloroplasts exhibited the fastest and the most obvious reaction to freeze-thaw damage.Several sharply freeze-thaw cycles resulted in damage to plasma membranes,severe damage to organelles,dehydration of cells and cell death.
Discussion
Southern P.deltoides cultivars do not survive the winter in Liaoning Province,even in years with normal weather conditions.Therefore,poplar distribution is limited by temperature.Tao and Jin(2005)and Sakai and Larcher(1987)hypothesized that under conditions of cooling-induced extracellular ice,different plant materials have different lethal temperature ranges determined by frost resistance.When the absolute low temperature in winter is lower than the minimum frost resistance of poplar,the poplar dies.It is widely accepted that low temperatures are the factor limiting tree distribution.
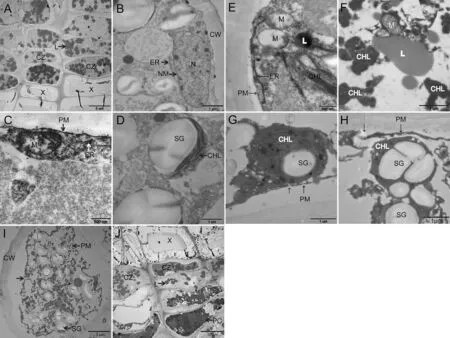
Fig.2 Test results of ultrastructure.a-d Cell that was not frozen and thawed.a Shows the phloem PC and CZ.The chloroplasts were present in phloem PC.Cambium and xylem cells had no chloroplast.The radial axis of cambium cells was short,and the tangential axis was long.The cell wall was thin,containing vacuoles of unequal size.b The nucleus,ER and other organelles of the cambium cells were squeezed against the edge of the cell wall by the vacuole;the double nuclear membrane was clear,and the rough ER and L were visible.c,d show the phloem PC.The mitochondrial cristae of C were clearly visible.The PM was intact,and the ER was clear.d The CHL contained starch grains,and the grana lamellae were clear.Black punctate osmiophilic granules were visible.e-h PC after one freezethaw treatment.The cytoplasm and CW were separated in E.The mitochondria were hollow.The CHL membranes were degraded,and the lamellar structure was disrupted,but the ER was normal,which indicated that CHL and M respond to freezing and thawing before the ER does.The M in f was normal,but the CHL membrane was degraded and the grana lamellae were completely dispersed,which showed that the CHL responds to freezing and thawing before the M does.The CHL membranes in g,h were completely degraded,and the PM was damaged.i-j Cell of the cambium and phloem after four freeze-thaw treatments.The organelles in i were destroyed and no longer discernable;In j,the cambium and phloem PC show severe plasmolysis,PM degradation and damage,cell dehydration and death.ER Endoplasmic reticulum, CHL chloroplast, M mitochondrion,N cell nucleus,NM nuclear membrane,CW cell wall,PM plasma membrane,SG starch grain,P phloem,X xylem,CZ cambium zone,L lipid droplet,PC parenchyma cell
The frost resistance of trees is changing(Martz et al.2006), and frost resistance differs by tree species. In addition,the frost resistance of a given tree species differs by age,season,and tissue.Frost resistance is lowest in dormant trees.When flowering dogwood specimens were cooled from 0 to-60°C at a rate of 5°C h-1,intracellular ice formation was observed at -20, -10 and-5°C in the winter,spring and summer,respectively(Ristic and Ashworth 1993b).Therefore,although freezing injury does not occur in poplar during dormancy,poplar clones can still sustain freezing injury at other times.Frost resistance is greater in poplar during cold acclimation and de-acclimation than during dormancy;during this time,when the temperature suddenly drops, freezing injury occurs.Cold spells in the late spring usually produce this type of freezing damage.In March 2009-2010,however,the temperature reached its lowest point(-16.3°C)of the 5-year period,but winter deaths in poplar did not occur during this year.This result demonstrates that low temperatures are not sufficient to cause overwintering death as one occurrence of low temperatures will cause frost damage but will not necessarily kill overwintering poplars.
It is generally thought that the degree and duration of low temperatures are two important limiting factors for poplar distribution(Lyons 2003).Our data showed,however,that when poplar clones that can overwinter in normal years encounter low temperatures or low negative accumulated temperatures(cold temperatures for a long duration),they do not necessarily die in the winter.Stem frostcrackingrate,large-tree DR,and seedling DR were not correlated with annual negative accumulated temperature,winter minimum temperature,or winter average temperature.This demonstrates that the cause of poplar overwintering death is not long-term exposure to low temperatures.In general,trees do not die after exposure to freezing temperatures for two reasons:the presence of extracellular ice and the depth of supercooling and vitrification(Strimbeck et al.2008).Xylem parenchyma cells of apple trees commonly exploit supercooling as a strategy to avoid frost damage(Sakai and Larcher 1987).This strategy has been reported for many temperate trees(Burke et al.1976).The extent of supercooling is limited to approximately-40°C due to the homogeneous nucleation temperature of water(Malone and Ashworth 1991).In the current study,the minimum temperature employed for the freeze-thaw cycle test was therefore-40°C,sufficient to ensure frost formation and occasionally recorded in nature.
Field observations revealed that the fluctuation in the temperature of the tree body was significantly greater than the change in air temperature.As seen in poplar sunlightexposed bark in February 2009,which had the largest diurnal range(up to 38.6°C),intense freeze-thaw cycles can have a considerable impact on poplar overwintering.Laboratory tests showed that exposure to drastic negative and positive fluctuations in temperature around 0°C caused more damage than exposure to constant low temperatures.Resistance to freeze-thaw cycles differed among clones(Table 3);cold-resistant clones had low relative electrical conductivity values.As a general rule,the more freeze-thaw cycles,the lower the bud-burst rate;these cycles heavily damaged poplar branches.Each clone has some resistance to freeze-thaw cycles.Although the winter of 2008-2009 was relatively warm, the overwintering death of poplar occurred overlarge areas;thus,overwintering death may be associated with exposure to freezethaw cycles.
Our laboratory tests revealed that the bud-burst rates of poplar clones with different levels of cold hardiness were similar(P >0.05)after one freeze-thaw cycle,with budburst rates of 90% or more. Ultrastructural analysis revealed that after a freeze-thaw cycle,chloroplasts were the main organelles that were damaged,and this damage was reversible.This result demonstrates that damage to trees is relatively minor after a freeze-thaw cycle.Lund and Livingston(1999)examined changes in chlorophyll absorbency in red spruce(Picea rubens Sarg.)foliage in response to simulated freezing cycles. No detectable chlorophyll degradation occurred after one treatment cycle in any trees on any sampling date.
The cambium is key to tree survival.Differences were observed in the degree of damage to cambium ultrastructure after different freeze-thaw cycles in the same poplar clones. We observed that the chloroplast reacts most strongly and quickly to freeze-thaw damage,followed by the mitochondria and endoplasmic reticulum. Poplar phloem and cambium exhibited little ultrastructural damage after one freeze-thaw cycle.After several freeze-thaw cycles,when the accumulation of freeze-thaw damage exceeded the tolerance of the cells,the plasma membrane,chloroplast, mitochondria, endoplasmic reticulum and other organelles degraded and the cell died due to loss of function and dehydration.
Based on the above analysis,we suggest the following three definitions:(1)freezing tolerance degree is the lowest temperature that tissues or cells can endure for a certain period of time and not exhibit irreversible damage;(2)effective freeze-thaw is the process in which cells or tissues freeze and then thaw completely-the cells or tissues suffer irreversible damage after exposure to temperatures lower than the freezing tolerance degree;(3)freeze-thaw resistance degree is the maximum number of effective freeze-thaw cycles that cells or tissues can undergo without dying in a certain period of time.
We also propose a cumulative-effect hypothesis describing effective freeze-thaw damage:(1)exposure to low temperatures is necessary but not sufficient for the occurrence of overwintering death in trees;(2)the freezing tolerance degree is greater than or equal to the frost resistance;(3)cells or tissues suffer irreversible damage from temperatures equal to or lower than the freezing tolerance degree;(4)effective freeze-thaw cycles often cause the accumulation of irreversible damage to cells or tissues-when the accumulation of irreversible damage is greater than the freeze-thaw resistance degree of tree tissues or cells,the tree tissues or cells die;(5)the cause of cell death is severe damage to organelles,degradation of the plasma membrane and other cell parts,dehydration of cells and subsequent loss of organelle function.
The difference between this hypothesis and the frostresistance and freezing-injury hypothesis is as follows:this hypothesis emphasizes the cumulative effect of effective freeze-thaw damage:tree overwintering death involves not only low temperatures but melting;one freezing event is not necessarily fatal,and the cumulative effect of many effective freeze-thaw cycles can also lead to tree death.This hypothesis does not deny the existence of frost resistance but,rather,takes freezing injury into account.
In addition to the results of the current study,other evidence supports the current hypothesis:(1)Frost damage in plants includes both reversible and irreversible damage.The plasma membrane,which is considered the original location of freezing injury,exhibits four different degrees of freezing damage(Iswari and Palta 1989):slight(reversible),partial(partially reversible),substantial and total(irreversible).Freezing-induced osmotic contraction results in endocytotic vesiculation of the plasma membrane,which is irreversible(Webb et al.1994).(2)The membrane lipid phase transformation theory(Lyons 2003;Steponkus 1984;Steponkus and Wiest 1978)suggests that membrane lipid phase transformation induced by temperature conditions is completely reversible;thus,exposure to low temperatures for a short period of time does not cause irreversible damage.Irreversible damage occurs only when the duration of low-temperature treatment is long enough to induce degenerative changes throughout the entire system.(3)The plasma membrane ATP enzyme inactivation theory(Hellergren et al.1987)and the free radical theory(Asada and Kanematsu 1976; Heber 1967) suggest that H+-ATP enzyme inactivation in the plasma membrane and the membrane degreasing effect are reversible in the early stage.When freezing stress continues,the free radical scavenging system is impaired and the damage becomes irreversible,eventually leading to cell death.
In overwintering poplar,successive strong fluctuations in temperature can lead to numerous freeze-thaw events,especially in bright sunlight conditions,where increases in the temperature of southwest-facing near-surface bark tissue caused by sunlight and ground radiation lead to partial tissue dehardening and thus reduced frost resistance.The temperature decreases at night,and the tree freezes and then thaws in the daytime,forming a effective freeze-thaw cycle(which can occur over the course of one or several days).When the accumulated damage produced by repeated effective freeze-thaw cycles exceeds the freeze-thaw resistance degree of the cambium,the poplar dies.Splitting section-type injuries occur on the southwest-facing surface,a phenomenon closely related to sunlight exposure.Short periods of defrosting under warm conditions are the main risk to poplar,and the accumulation of effective freezethaw damage over the limit of freeze-thaw resistance degree is the main reason for poplar winter death.Most winter death in poplar appears to involve dehardening,whereas temperature changes during the dormant season have little effect on poplar wintering.
Due to global warming,the cultivation limit of fastgrowing black poplar is moving northward.Survival may be threatened when,in a changing climate,frost hardening and dehardening occur at the wrong time(Ha¨nninen and Tanino 2011),particularly when temperature is the predominant controlling factor for these processes(Heide and Prestrud,2005).Sakai and Larcher(1987)determined that plant survival in the winter is the result of relatively complex interactions,many of which have still not been described quantitatively or qualitatively.Further research is needed to elucidate the mechanism of winter death in poplar.
AcknowledgementsWe thank the College of Biological Sciences,China Agricultural University,for technical support in the ultrastructure test.
Author contributions(1) CY made substantial contributions to conception and design,conducted tests,and drafted the article;(2)JP participated in the acquisition of data and data analysis;(3)XL participated in the analysis and interpretation of the data;(4)DL participated in the acquisition and analysis of the data; (5) ZY participated in the acquisition and analysis of the data;(6)YZ participated in the acquisition and analysis of the data.
Compliance with ethical standards
Conflict of interestThe author declares that they have no conflict of interest.
杂志排行

Journal of Forestry Research的其它文章
- Protective and defensive roles of non-glandular trichomes against multiple stresses: structure-function coordination
- Assessment of early survival and growth of planted Scots pine(Pinus sylvestris)seedlings under extreme continental climate conditions of northern Mongolia
- Influencing in vitro clonal propagation of Chonemorpha fragrans(moon)Alston by culture media strength,plant growth regulators,carbon source and photo periodic incubation
- Variation analysis of growth traits of four poplar clones under different water and fertilizer management
- Nodule study in Albizia chinensis in relation to nitrogen metabolism,morphology and biomass
- Comparative transcriptome analyses reveal candidate genes regulating wood quality in Japanese larch(Larix kaempferi)