长链非编码RNA与Wnt/β-catenin通路相互作用在消化道肿瘤中的研究进展
2020-01-17陆佳伟张文波蒋鹏程
陆佳伟 张文波 蒋鹏程
江苏大学附属人民医院普外科(江苏镇江212002)
据估计,2018年全球有1 810 万例新诊断癌症患者和960 万名患者死亡,与英国和美国相比,中国的癌症发病率较低但死亡率却较高,其中约36.4%的患者死于消化道肿瘤(胃癌、肝癌和食道癌),而在英国或美国死于消化道肿瘤的患者占总癌症死亡率不到5%,造成这种现状的原因在很大程度上是由于早期癌症的诊断率较低[1]。目前,消化道肿瘤的早期诊断主要依赖于血清学、影像学、内镜等检查,但由于特异性较低导致消化道肿瘤在被发现时往往已经处于晚期,相关的新辅助放射治疗、辅助化疗、外科手术等治疗手段并不能显著改善患者的预后。长链非编码RNA(long non-coding RNA,lncRNA)曾一度被认为是“转录噪音”,但近年来的相关研究表明,其在人类肿瘤的发生和发展中起着重要作用[2]。研究[3]证实,lncRNA能够通过参与调控多种信号通路,如Wnt通路、p53通路和NF-κB通路等,从而调控肿瘤细胞增殖、迁移等能力。其中,Wnt/β-catenin是肿瘤发生发展的关键信号,能够调控肿瘤细胞的生长、衰老、凋亡、分化和转移等诸多过程[4]。关于lncRNA 与Wnt/β-catenin 通路相互作用的研究是消化道肿瘤基础研究领域的热点问题,但目前尚缺乏归纳总结。本文就相应问题的最新研究展开综述,以期为消化系统肿瘤预测、诊断和治疗的研究提供新的思路。
1 LncRNA 与消化道肿瘤的关系
LncRNA是一组长度大于200 nt的非编码RNA,主要在表观遗传、转录、转录后和翻译水平调控基因表达[5-6]。在许多癌症中lncRNA 的表达异常与癌症的进展、转移和侵袭等紧密相关。因此,lncRNA 有潜力成为诊断癌症的生物标志物[7]。LncRNA 常通过不同的信号通路影响着消化道肿瘤的发生发展,其功能主要可以分为两类:促癌lncRNA 和抑癌lncRNA。例如lncRNA-NR027113在胃癌患者组织中过表达并且能够通过激活上皮细胞-间充质转化(epithelial-mesenchymal transition,EMT)通路促进胃癌细胞的迁移和侵袭[8]。LINC00472 在肝癌组织表达降低,作为一种抑癌基因能够通过miR-93-5p/PDCD4 通路抑制肝癌细胞的增殖、迁移和侵袭,促进癌细胞凋亡[9]。目前,lncRNA 在消化道肿瘤如何调控这些通路仍未明确,但随着研究的进展,这些问题有望得到解决并应用于消化道肿瘤的诊断和治疗中。
2 Wnt/β-catenin 通路和消化道肿瘤之间的关系
Wnt/β-catenin 通路是一种研究得比较多的Wnt 经典通路,该通路主要通过调节转录共激活因子β-catenin 的活化,进而控制通路下游基因的表达来发挥作用[10]。当没有Wnt 配体存在时,细胞质β-catenin 与轴蛋白(Axin)、肿瘤抑制性腺瘤息肉病基因产物(APC)、糖原合成酶激酶3(GSK3)和酪蛋白激酶1(CK1)形成复合物,并被CK1 和GSK3 磷酸化,而后磷酸化的β-catenin 由E3 泛素连接酶β-Trcp 蛋白识别并泛素化,泛素化后的β-catenin 最终被蛋白酶体降解[11]。当Wnt 配体和细胞膜上低密度脂蛋白受体相关蛋白(LRP5/6)和卷曲蛋白(FZD)结合时,经典Wnt通路被激活,触发细胞内的信号转导,活化细胞质蓬乱蛋白Dhs 或Dvl,进而抑制细胞质内β-catenin 与Axi、GSK-3β、CK1、APC 等形成降解复合物[12]。随后,β-catenin在细胞质中稳定下来并且积聚,穿过核膜进入细胞核并与T 细胞转录因子/淋巴样增强因子(TCF/LEF)相互作用,组成复合物并相应地转录激活CyclinD1、c-Myc、CD44 等下游靶基因[13-14]。
研究表明,Wnt/β-catenin 通路的异常激活与消化道肿瘤的发生发展密切相关,其中以结直肠癌联系最密切,约80%的结直肠癌患者有APC基因位点的突变,10%的患者在β-catenin 上发生突变[15]。不少文献表明异常激活的Wnt/β-catenin 通路对于消化道肿瘤的EMT、抗肿瘤药的耐药性、肿瘤干细胞自我更新能力等方面产生重要影响。比如,STEMMER 等[16]发现Wnt/β-catenin 通路激活可以使Slug、Snail、Twist 等表达增加,降低E-cadherin表达,促进EMT 形成。在结直肠癌患者中Wnt/βcatenin 信号通路异常激活后会导致患者放化疗耐药性增高,相关肿瘤细胞凋亡减少[17]。此外,Wnt/β-catenin 通路在肝癌患者中还能调控肝正常或肿瘤干细胞的自我更新能力[18]。
3 LncRNA 和Wnt/β-catenin 通路的相互作用对消化道肿瘤的影响
3.1 食管癌(esophageal cancer,EC) EC 是一种高发的消化道肿瘤,在所有癌症中死亡率排第六位[19]。因此,加强对高发人群的早期筛查对于改善患者预后有着积极意义。通过研究EC 的发病机制有助于寻找潜在的特异性较高的新的肿瘤标志物,提高早期诊断率。研究表明lncRNA 的表达及Wnt/β-catenin 通路在EC 患者中存在异常,从而影响EC 的生长、分化等过程。LINC00675 在EC 组织中低表达,与EC 的淋巴结转移程度、肿瘤分期等紧密相关,在体外实验中当LINC00675过表达时,可以通过抑制Wnt/β-catenin 通路进而抑制EC 细胞的增殖、迁移、EMT 等方面[20]。LncRNA FEZF1-AS1 在EC 表达增高,沉默FEZF1-AS1 时能够明显抑制EC 细胞的迁移和侵袭能力。进一步研究发现EC 组织中β-catenin 表达增高,当沉默FEZF1-AS1 时会抑制其表达,反之促进,表明lncRNAFEZF1-AS1 能够通过Wnt/β-catenin信号通路促进EC 细胞迁移和侵袭[21]。LncRNA DUXAP8在EC 组织表达增高,敲低DUXAP8 后β-catenin、Cyclin D1和c-Myc表达下降[22]。还有一些lncRNA可以间接调节Wnt/β-catenin 通路。比如,HOTAIR 在EC 组织高表达,能够通过促进Wnt 通路抑制因子1(WIF-1)启动子区域组蛋白H3K27 的甲基化从而下调WIF-1,激活Wnt/β-catenin 通路[23]。LncRNA MEG3 在EC 表达下降,作为内源性竞争RNA 与mir-4261 结合上调Wnt/β-catenin 通路的抑制因子DKK2 从而抑制Wnt/β-catenin 通路[24]。目前,在EC 中Wnt/β-catenin 通路对lncRNA 的调控报道尚少。总之,在EC中lncRNA 通过Wnt/β-catenin 通路参与调控EC 细胞的增殖、迁移、侵袭、EMT 等过程,相关机制的进一步研究可能有助于更好地诊断和治疗EC。
3.2 胃癌(gastric carcinoma,GC) GC 是全球第四大最常见的恶性肿瘤,全球发病率呈上升趋势[25]。然而大多数GC 患者由于缺乏典型的早期症状而延误诊断。LncRNA 与Wnt/β-catenin 信号通路相互作用可以调控GC 的进展。一方面,一些lncRNA 可以通过直接或间接的方式调节Wnt 通路而抑制GC 的发展。如LINC01133 在GC 组织低表达,作为miR-106a-3p 的内源性竞争RNA 抑制miR-106a-3p 表达,促进APC 基因表达增高从而减少β-catenin 在细胞核的积聚,进而抑制EMT和转移[26]。类似地,抑癌基因LINC01314 过表达时能够下调KLK4 从而导致Wnt/β-catenin 等表达下降,抑制GC 细胞的迁移、侵袭和肿瘤血管生成[27]。另一方面,不少lncRNA 在GC 组织中表达水平增高,并通过激活Wnt/β-catenin 通路促进GC细胞增殖和转移。例如,LINC00052 可以通过与β-catenin 和甲基转移酶SMYD2 相互作用,使β-catenin 发生甲基化以维持其稳定性,激活Wnt/β-catenin 通路[28]。此外,HOTAIR 在GC 组织中表达水平增加,研究表明其与mir-34a 竞争性地结合。当HOTAIR 被敲低时mir-34a 表达增高进而通过抑制PI3K/Akt 和Wnt/β-catenin 通路,减少GC 细胞对顺铂的耐药性[29]。因此,lncRNA 可以通过Wnt/β-catenin 通路影响GC 细胞的迁移、侵袭、耐药性、EMT 和肿瘤转移等方面。不仅如此,GC 中一些lncRNA 也可以被Wnt/β-catenin 通路下游靶基因c-Myc 所调控。例如,c-Myc 可以诱导lncRNA H19 在GC 组织中表达增高从而影响肿瘤细胞的增殖[30]。类似地,c-Myc可以直接与lncRNA CCAT1启动子区域的E-box 元件相结合上调CCAT1 从而促进癌细胞的增殖和迁移[31]。
3.3 结直肠癌(colorectal cancer,CRC) 在中国CRC 的发病率已上升至第三位,仅次于肺癌和胃癌,死亡率位居第五位[32]。因此,寻找CRC 的早期诊断生物标志物及治疗靶点有望提高患者的生存率。LncRNA 可以通过调控Wnt/β-catenin 通路影响CRC 的发生发展,如转录因子YY1 可以转录激活ARAP1-AS1 进而激活Wnt/β-catenin 通路而促进CRC 的迁移、侵袭和EMT[33]。LncRNA GAS5在肠癌组织低表达,当其上调时可以抑制Wnt/β-catenin 通路进而抑制肠癌的肿瘤血管生成、侵袭和肝转移等[34]。Wnt/β-catenin 通路也可以反过来调节一些lncRNA 的表达,如Wnt/β-catenin 通路的异常激活可以促进其靶基因c-Myc 的转录激活从而上调lncRNA MYU,而MYU 与RNA 结合蛋白hnRNP-K 的结合能维持CDK6 的表达,促使细胞周期发生G1-S 转换,从而调节结肠癌细胞的增殖和肿瘤形成[35]。SUNAMURA 等[36]发现在CRC 中当细胞核中的β-catenin 累积增多时,lncRNA KCNQ1OT1 转录水平相应增高。当敲低β-catenin 时lncRNAKCNQ1OT1 转录水平也随之下降,这说明β-catenin 可能通过直接调节KCNQ1OT1 促进CRC的进展。不仅如此,lncRNA 与Wnt/β-catenin 通路形成的反馈调节环能够调节CRC 增殖、侵袭和迁移。例如,XIE 等[37]运用GEO 数据库分析敲低β-catenin 后差异表达的lncRNA,筛选出高表达的lncRNA BCAT1。进一步的研究发现BCAT1 过表达时能下调Wnt/β-catenin 信号通路的靶基因Cyclin D1、c-Myc 和MMP-2 等,进而抑制CRC 细胞的增殖和转移。此外,对lncRNA 与CRC 患者化疗耐药性和放疗敏感性关系的研究发现,lncRNA H19 在甲氨蝶呤耐药的癌株中表达增高,当H19 被敲低后CRC 对甲氨蝶呤敏感性增加,反之降低。研究其机制发现H19 通过激活Wnt/β-catenin 信号通路介导甲氨蝶呤的耐药[38]。WANG 等[39]发现lincRNAp21 通过调控Wnt/β-catenin 通路增加放疗敏感性,促进细胞凋亡。总之,CRC中lncRNA与Wnt/β-catenin 通路相互作用能影响肿瘤形成、增殖、肝转移、肿瘤血管生成、放化疗敏感性和EMT等。
3.4 胰腺癌(pancreatic cancer,PC) PC 是一种恶性度极高的消化系统肿瘤,其早期症状隐匿,发现时多已是晚期,手术切除率低,预后极差。弄清PC 相关病理机制有助于寻找有效的靶点进行针对性的治疗。近年来,有研究表明lncRNA 与Wnt/β-catenin 通路相互作用能够影响PC 进展。LncRNA DLX6-AS1 在PC 组织和细胞株中表达增高,过表达的DLX6-AS1 能促进PC 细胞的增殖、迁移和侵袭并抑制细胞凋亡。进一步研究发现,DLX6-AS1 与miR-497-5p 结合时能逆转其对FZD4、FZD6 的抑制作用,从而上调β-catenin 促进PC的发生[40]。在转录因子FOXO1 高表达的PC 细胞中,LINC01197 作为抑癌基因在PC 组织表达水平明显下降。从机制上,LINC01197 能够在细胞核中与β-catenin 相结合阻止其与转录因子TCF4 结合进而抑制Wnt/β-catenin 通路表达,抑制PC 的发生发展[41]。JIANG 等[42]的研究表明,lncRNA HOTAIR在PC 中表达增高,当敲低HOTAIR 时WIF-1 表达增高进而抑制Wnt/β-catenin 通路表达导致PC 的放疗敏感性增高。此外,lncRNA HOTTIP 在PC 表达增高,可以与WDR5 结合上调HOXA9 激活Wnt/β-catenin 通路促进胰腺肿瘤干细胞的形成[43]。不仅如此,YU 等[44]研究发现Wnt/β-catenin 通路下游靶基因c-Myc 与CCAT1启动子区域的E-box 元件结合而激活CCAT1,进而影响PC 肿瘤的生成和转移。由此可见,在PC 中lncRNA 与Wnt/β-catenin 的相互作用可以影响肿瘤细胞的增殖、转移、侵袭、放疗敏感性和肿瘤干细胞生长分化等。
3.5 肝癌(hepatocellular carcinoma,HCC) 近年来许多研究表明lncRNA 可以作用于Wnt/β-catenin通路中的不同组分从而调控HCC 的发生发展。LncRNA DGCR5在HCC患者中表达下降,当DGCR5过表达时能够抑制HCC 细胞的生长、迁移和侵袭。进一步研究表明,DGCR5 通过下调β-catenin、CyclinD1 和上调GSK-3β抑制Wnt/β-catenin 通路激活进而抑制HCC 的进展[45]。LncRNA-LALR1 通过将CTCF 导入Axin1 的启动子以阻止Axin1 转录,抑制含APC 的降解复合物的形成从而激活Wnt/β-catenin 通路[46]。LncTCF7 在HCC 表达水平增高,通过将SWI/SNF 复合物导入TCF7 的启动子区域进而激活Wnt 通路,促进人类肝肿瘤干细胞的自我更新[47]。不仅如此,Wnt/β-catenin通路也可以调节一些lncRNA 表达。例如,有研究发现在肝肿瘤细胞中Wnt/β-catenin通路激活时可以上调lncRNA uc.158。进一步的体外实验表明在CTNNB1 突变的HCC 细胞中,uc.158 表达水平较未突变的细胞表达水平增高,即通过调控β-catenin 表达能够改变人肝肿瘤细胞中的uc.158 表达水平[48]。ZHU 等[49]发现Wnt 通路的下游靶基因c-Myc 可直接结合CCAT 启动子区中的E-box 元件,并且当其异位表达时能增加启动子活性和CCAT1 的表达。此外,与高表达组相比,CCAT1 低表达的患者表现出更好的总体和无复发存活率。总之,在HCC 中lncRNA 可以与Wnt/β-catenin 通路的相互作用以调控肝肿瘤细胞的生长和分化。本文总结了Wnt/β-catenin通路相关的lncRNAs在消化道肿瘤中作用,见表1。
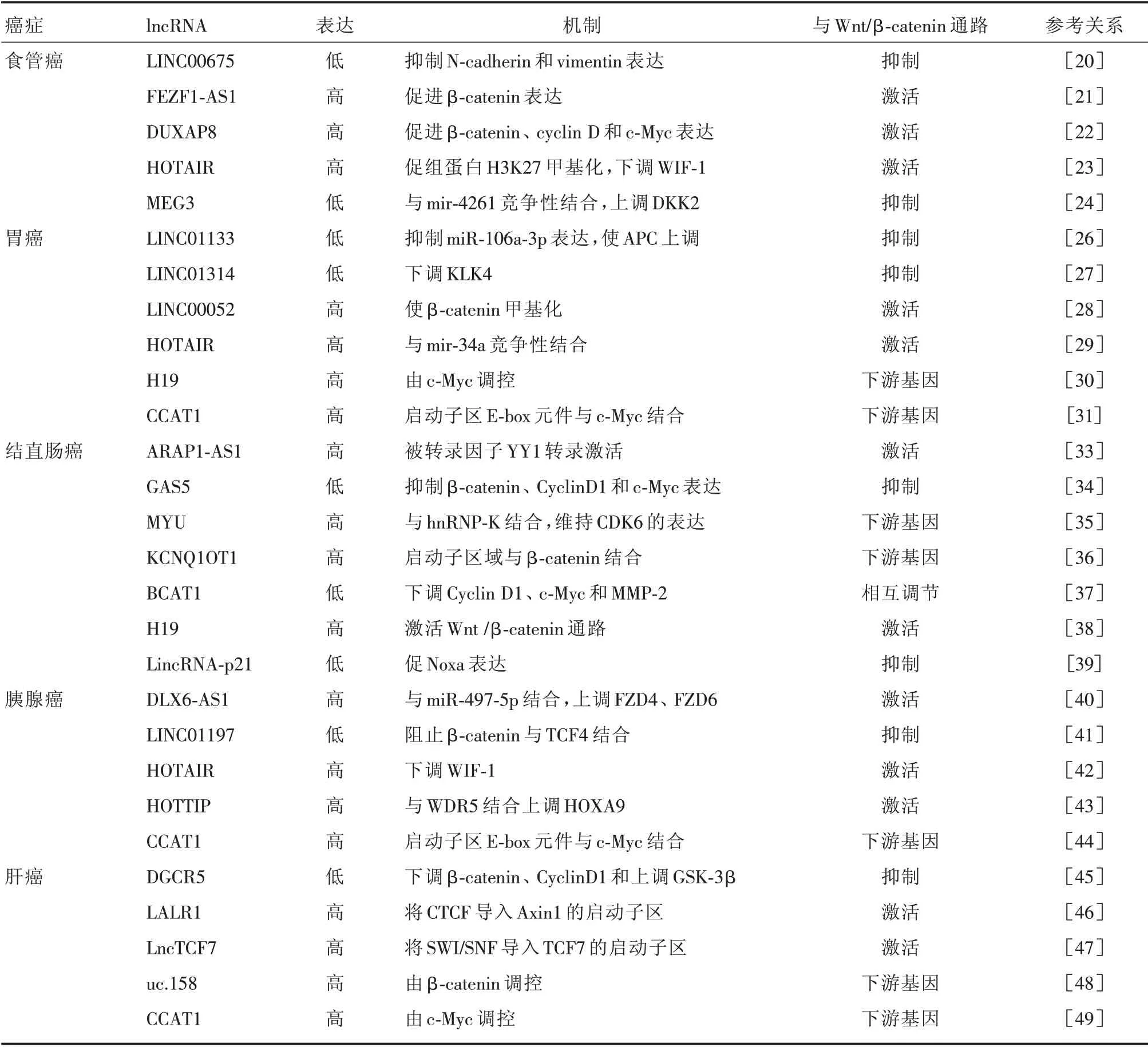
表1 Wnt/β-catenin 通路相关的lncRNAs 在消化道肿瘤作用Tab.1 Roles of Wnt/β-catenin pathway-related lncRNAs in gastrointestinal cancers
4 总结与展望
本文综述了近年来lncRNA 与Wnt/β-catenin通路相互作用在消化道肿瘤中的研究进展。这对研究消化道肿瘤中lncRNA 及Wnt/β-catenin 信号通路相关分子能否作为个体化治疗的潜在靶点有一定的意义。在消化道肿瘤中,lncRNA 作为Wnt/β-catenin 通路下游因子常被c-Myc 等分子所调控,c-Myc 如何调控这些lncRNA 有待深入研究。另一方面,lncRNA 对Wnt/β-catenin 通路的作用机制需进一步探讨。相同的lncRNA 在不同消化道肿瘤中可以不同方式参与调控Wnt/β-catenin 通路从而发挥不同作用。比如,HOTAIR 在胰腺癌和胃癌激活Wnt/β-catenin 通路的机制不同产生了不同的影响,需深入研究造成这种差异的因素是什么。此外,在不同消化道肿瘤中,不同的lncRNA 可通过相似的机制影响Wnt/β-catenin 信号通路,比如很多lncRNA 均可直接调控β-catenin 表达影响相关肿瘤的增殖和凋亡,这些lncRNA 之间是否存在协同作用需要进一步的研究。随着研究的不断深入,lncRNA 及Wnt/β-catenin 通路相关分子有望成为消化道肿瘤的早期诊断标志物并为消化道肿瘤的治疗提供有效的治疗靶点。
