魔芋葡甘聚糖对冷冻小麦面团面筋蛋白结构和功能特性的影响
2020-01-07郭金英贺亿杰韩四海任国艳罗登林刘建学
郭金英,贺亿杰,韩四海,任国艳,吴 影,罗登林,刘建学,2
(1.河南科技大学食品与生物工程学院,河南 洛阳 471023;2.河南省食品原料工程技术研究中心,河南 洛阳 471023;3.食品加工与发全国家级实验教学示范中心,河南 洛阳 471023)
冷冻面团技术主要指在面制食品生产过程中,运用冷冻冷藏原理和方法处理半成品或成品,待需用时经解冻处理,继而加工成为成品[1]。对于普通的面制食品,小麦面筋蛋白通过赋予面团吸水能力、内聚力和黏弹性,在面团强度、延展性和气体滞留性方面起着决定性作用,其质量也直接决定了面团的最终食用品质[2]。而在冷冻过程中,冰重结晶和二硫键(S—S)断裂诱发面筋蛋白高分子质量亚基解聚,破坏面筋蛋白骨架结构,对面筋蛋白造成机械性能损失。可使用亲水胶体、乳化剂和其他面团改良剂减轻冷冻对面筋蛋白的影响。亲水胶体良好的亲水持水特性减少了游离态水,提高了冷冻食品体系的冻融稳定性,胶体能够控制面团中冰结晶的生长速率和冰结晶的大小,抑制面筋网络结构的破坏[3]。亲水胶体中因其某些基团能够与面团中淀粉及蛋白质作用,改变面团中淀粉和蛋白质特性,范鹏辉[4]在冷冻面团配方中添加黄原胶增加面团的筋力、吸水率和持气性,从而改善冷冻对面筋蛋白的机械损伤。胶体能够与面筋蛋白相互作用,形成胶体-面筋蛋白复合物,增大面筋蛋白链的长度,强化面筋网络,提高面筋结构稳定性,如黄原胶[5]和卡拉胶[6-7];也有胶体自身形成网络,加固面团的面筋蛋白网络结构,如魔芋胶能够形成糖网络[8]。
魔芋属于天南星科魔芋属植物,含有丰富的魔芋葡甘聚糖(konjac glucomannan,KGM),KGM是由葡萄糖和甘露糖以β-(1,4)糖苷键连接的高分子杂多糖,是众所周知的可溶性水胶体[9]。同时,它也是一种优良的膳食纤维,具有清洁肠道、提高糖耐量、预防肥胖和改善胆固醇代谢等功能[10],具有高保水性、膨胀性、增稠性、胶凝性、乳化性、悬浮性和黏结性,其性能优于普通食品凝胶,如琼脂、角叉菜胶和明胶等,因而越来越多地被用作食品添加剂[11-13]。当KGM添加到面团中时,可以显著增加面团的吸水性和发酵时间,同时通过增强面筋网络影响面团的硬度和弹性,在低蛋白小麦面粉中产生更紧凑和连续的微观结构[14]。在冷冻食品中,KGM已被用于冷冻饺子中以防止霜冻开裂并延长冷冻饺子的保质期[8]。同时,KGM还用于改善冷冻面团的流变性、微观结构和冷冻稳定性[15]。
当前,对KGM研究主要集中在新鲜面团中作为复合添加剂方面,而关于KGM单独作为添加剂应用于冷冻面团的研究较少,对冷冻面团面筋蛋白的结构和功能特性影响研究则更少。因此本实验从小麦面粉中分离得到面筋蛋白质,研究添加KGM对面筋蛋白结构和功能性质以及微观物理结构的影响,为KGM在冷冻面团中的应用提供理论依据。
1 材料与方法
1.1 材料与试剂
魔芋精粉(纯度95.10%) 成都路特食品有限公司;面粉(蛋白质质量分数11.63%,含水率13.70%) 河南省驻马店市一加一天然面粉有限公司;三羟甲基氨基甲烷(Tris (hydroxymethyl) methyl aminomethane,Tris)美国Sigma公司;甘氨酸 上海蓝季科技发展有限公司;溴酚蓝、考马斯亮蓝R-250 天津市旭泰化学试剂有限公司;四甲基乙二胺 国药集团化学试剂有限公司;蛋白质标准品 宝生物工程有限公司。
1.2 仪器与设备
HM740型和面机 青岛汉尚电器有限公司;RE-52A旋转蒸发仪 巩义市瑞德仪器设备有限公司;DSC-1型差示扫描量热(differential scanning calorimetry,DSC)仪 瑞士梅特勒托利多仪器有限公司;NMI20-015V-1核磁共振成像分析仪 上海纽迈电子科技有限公司;UV-4800型紫外-可见分光光度计 尤尼科科学仪器有限公司;VERTEX70型红外光谱仪 德国布鲁克光谱仪器公司;LGJ-10D冷冻干燥机 北京四环科学仪器厂有限公司;TM3000日立台式电镜 日立高新技术公司;DHR2流变仪 美国TA仪器有限公司。
1.3 方法
1.3.1 样品制备
参考Graberger等[16]面粉脱脂的方法,略有改进。面粉和三氯甲烷室温下按照1∶2(g/mL)的比例混合,搅拌5 min进行脱脂、抽滤,再重复2 次,时间分别为1 h和1.5 h。脱脂面粉于室温通风橱中晾干,直至闻不到氯仿气味后过200 目筛。取400 g脱脂面粉,加入250 mL 0.4 mol/L的NaCl溶液,在搅面机里混合5 min,静置20 min促使面筋形成,用NaCl溶液反复手洗面团直至将淀粉完全除去,得到湿面筋。将上述湿面筋样品用蒸馏水洗涤除去残留的NaCl溶液,冷冻干燥后粉碎,过200 目筛,即得面筋蛋白粉[17]。
将2 g冻干面筋蛋白粉末分散在含有质量分数为1%十二烷基硫酸钠(sodium dodecyl sulfate,SDS)的100 mL磷酸盐缓冲液中。称取面粉质量取代度为0%、0.5%、1.5%、2.5%、3.5%和4.5%的KGM溶于水并定容至100 mL。分别取10 mL KGM溶液和10 mL蛋白溶液配制成相应浓度的KGM-面筋蛋白,置于-70 ℃超低温冰箱中待中心温度降至-18 ℃,放置-18 ℃的冰箱中贮藏24 h,然后在4 ℃环境下解冻,解冻后的样品放在磁力搅拌器中,35 ℃搅拌2 h,以模拟其在面团制备中的反应。分为两部分:一部分用于液体样品实验测定;另一部分冷冻干燥并粉碎用于固体样品实验测定。
1.3.2 面筋蛋白持水性测定
在离心管中加入50 mg面筋蛋白,加入4 mL蒸馏水,旋涡混匀2 min,静置60 min,4 500 r/min离心10 min,除去上清液,称量面筋蛋白与离心管的总质量。同一样品重复测定3 次,计算加入不同水平的KGM后面筋蛋白持水性,如式(1)所示:
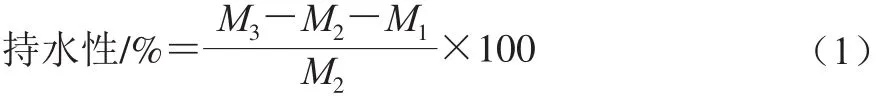
式中:M1为离心管质量;M2为面筋蛋白粉质量;M3为面筋蛋白与离心管的总质量。
1.3.3 面筋蛋白分子质量测定
根据Sotelo等[18]的方法稍作改进,进行SDS-聚丙烯酰胺凝胶电泳(SDS-polyacrylamide gel electrophoresis,SDS-PAGE)。使用5%浓缩凝胶和10%分离凝胶分析。将40 μL蛋白质溶液置于1.5 mL离心管中,加入40 μL上样缓冲液(含有20 mg/mL SDS,体积分数10%甘油的Tris-HCl,0.125 mol/L,pH 6.8)和质量分数0.01%的溴酚蓝)。样品沸水浴5 min,上样量为15 μL,在恒定电流100 A下直至蓝色染料迁移至凝胶末端附近,除去凝胶,进行染色、脱色直至凝胶背景清晰用于拍照。
1.3.4 面筋蛋白流变性测定
将解冻后不同水平的KGM-面筋蛋白样品分别放置在流变仪底板上,上板选用40 mm平板,平板之间间距设置为1 000 μm,在4 ℃进行动态频率扫描,应变设置为3%,扫描频率为1~10 Hz。
1.3.5 面筋蛋白热性质测定
根据Alberto等[19]DSC方法进行测定。准确称取2~3 mg冷冻干燥的面筋蛋白并密封在DSC的小铝锅中,然后在25~90 ℃范围,以5 ℃/min的恒定速率进行温度扫描,空锅用作参考。通过TA系统Muse软件计算变性峰值温度(Tp)和焓(ΔH)。
1.3.6 面筋蛋白游离巯基测定
将150 mg蛋白质样品悬浮于5.0 mL Tris-甘氨酸缓冲液(10.40 g Tris,480.00 g尿素,10.40 g甘氨酸,用质量分数1% SDS溶液稀释至1 000 mL)中,加入0.1 mL 5,5’-二硫双(2-硝基苯甲酸)(5,5’-dithiobis-(2-nitrobenzoic acid),DTNB)溶液(用质量分数为1% SDS溶液将40 mg DTNB调节至10 mL),10 000 r/min离心20 min。取上清液在412 nm波长处测定吸光度[20]。游离巯基含量计算如式(2)所示:

式中:A412nm为 412 nm波长处的吸光度;C为样品质量浓度/(mg/mL);5.02为稀释因子。
1.3.7 面筋蛋白二级结构测定
通过傅里叶变换红外光谱研究面筋蛋白二级结构。以4 cm-1的分辨率记录红外光谱,对于400~4 000 cm-1之间的每个样品扫描32 次。采用Omnic 8.2软件和Perkfit 4.0软件对酰胺I带进行去卷积处理,并求其二级导数得到特征峰,进行曲线拟合计算峰面积并确定面筋蛋白二级结构所占比例[21]。
1.3.8 扫描电子显微镜观察
将冷冻干燥后的不同浓度KGM-面筋蛋白,粉碎过200 目筛后,取样,用导电胶贴于置物台,置于扫描电子显微镜下,抽真空在真空环境下进行观察。
1.4 数据统计分析
Microsoft Excel、SPSS 17.0和Origin 8.5用于数据处理。通过单因素方差分析(ANOVA)和Duncan的多范围检验确定统计差异,P<0.05,差异显著。
2 结果与分析
2.1 KGM对面筋蛋白持水性的影响

图1 KGM取代度对面筋蛋白持水性的影响Fig. 1 Water-binding capacity of gluten with different contents of KGM
由图1可以看出,随着KGM取代度的增加,面筋蛋白持水量也不断增加,空白对照组的面筋蛋白持水性为2.01%,而KGM取代度0.5%的面筋蛋白持水性为3.95%,表明,KGM的添加显著增强了面筋蛋白的持水性。这主要由于KGM本身具有极强的吸水性,可以吸回自身质量80~100 倍的水分,所以KGM的添加必然会增加面筋蛋白的持水性[22]。面筋蛋白持水性的大小,间接反映了面筋蛋白内部网络结构均一性和稳定性[5]。随着KGM取代度的持续增加,面筋蛋白的持水性又分别增加了0.33%、0.73%、0.88%、1.54%。这可能因为KGM属于非离子型水溶性高分子胶体,其含有的丰富羟基与面筋蛋白相互作用,形成KGM-水-面筋蛋白复合网络结构,可以束缚更多的水分子,从而使复合体系持水性增强[23],使面筋蛋白结构更加稳定。持水性是蛋白质的重要功能性质,决定着蛋白质与食品体系中水分的相互作用[24]。
2.2 KGM对面筋蛋白分子质量的影响
面筋蛋白是自然界最为复杂的蛋白之一,分子质量可高达107Da。目前小麦面筋蛋白的分类主要沿用Osborne分离法,即根据面筋蛋白在乙醇水溶液中溶解度的差异,分为单体醇溶蛋白和聚合体谷蛋白,二者质量比接近1∶1[25]。面筋蛋白的SDS-PAGE图谱如图2所示,谷蛋白分为高分子质量谷蛋白(high molecular weight glutenin subunit,HMW-GS,约65~113 kDa)和低分子质量谷蛋白(low molecular weight glutenin subunit,LMW-GS,约30~60 kDa)。醇溶蛋白根据其在酸性pH值下的电泳迁移率分为4 组,分别为α-、β-、γ-和ω-醇溶蛋白。其中,α-和β-醇溶蛋白氨基酸和DNA非常相似,被称为“α型”醇溶蛋白。α和γ型为富硫亚基,分子质量在30~45 kDa之间。ω型为贫硫亚基,分子质量在46~74 kDa[26]。

图2 冷冻后面筋蛋白的SDS-PAGE分析Fig. 2 SDS-PAGE analysis of frozen gluten
图2 反映了不同KGM添加处理组的冷冻小麦面团面筋蛋白各亚基条带的变化,可看出,空白对照组的HMW-GS和LMW-GS条带清晰(泳道1),而所有样品处理组的条带颜色相对变浅(泳道2~6),这可能是由于KGM和面筋蛋白相互作用的结果。随着KGM取代度的增加,面筋蛋白条带随之加粗,颜色加深,更进一步证明KGM能够与面筋蛋白相互作用,且随着KGM取代度的增加,相互作用加强。空白对照组醇溶蛋白的ω型(泳道1)和所有样品处理组的面筋蛋白条带(泳道2~6)相比,并没有发现有任何条带发生移动现象。而空白对照组醇溶蛋白的α和γ型条带粗、颜色深(泳道1),所有样品处理组的面筋蛋白的条带较浅,颜色较淡(泳道2~6),但随着KGM取代度的增加,并未明显变化。在KGM存在下的低分子区域蛋白分子质量明显大于空白对照组,这可能是由于空白对照组原先聚集在浓缩胶的高分子质量蛋白发生解聚,小分子质量蛋白得以释放,从而使低分子质量蛋白浓度提高[27]。
2.3 KGM对面筋蛋白流变性的影响
从图3可看出,KGM-面筋蛋白黏弹特性参数随KGM取代度的不同而发生变化。与对照相比,随着KGM取代度的增加,面筋蛋白的储能模量(G’)和损耗模量(G〞)均呈增加趋势,而tanδ则呈下降趋势。G’是指经过一个振动周期的正弦形变后所恢复的能量,代表物质的弹性本质;G〞是指每个周期的正弦形变所消耗的能量,代表物质的黏性本质。面筋蛋白不仅具有黏性流体的一些特性,而且也具有弹性固体的一些特征,对面团的流变性起主要作用[28]。在相同的频率扫描下,当KGM取代度低于1.5%时,面筋蛋白的G’和G〞的增加均不明显;当KGM取代度高于1.5%时,面筋蛋白的G’和G〞均明显增加,表明KGM的添加使得KGM-面筋蛋白的弹性和黏性均得到提高。损耗角正切值tanδ的大小反映了体系中黏弹性的比例。从图3可知,在不同的频率扫描下,空白对照的tanδ呈逐步上升趋势;而在KGM存在下,面筋蛋白的tanδ值呈现先增加后降低的趋势,在KGM取代度为2.5%时,tanδ值达到最大。这表明2.5%的KGM的取代度对面筋蛋白弹性的贡献大于黏性,此时有利于谷蛋白分子之间形成交联结构,能够更好地赋予面团弹性,而当KGM含量增加,则更有利于面团黏性的增加。这主要是由于KGM是非离子型胶体,能够通过氢键和疏水键与面筋蛋白作用,并且位于支链上的官能团凝胶形成糖网络,强化面筋蛋白的网络结构,起到了保护面筋蛋白网络结构的作用,提高了冷冻面筋蛋白的冻藏稳定性[29]。

图3 KGM对面筋蛋白流变性的影响Fig. 3 Effect of KGM content on rheological properties of gluten
2.4 KGM对面筋蛋白热特性的影响

表1 KGM对面筋蛋白热特性的影响Table 1 Effect of KGM on thermal properties of gluten
由表1可知,与对照相比,所有样品处理组面筋蛋白变性温度(Tp)和变性焓(ΔH)均增高。随着KGM取代度的增加,相应浓度的KGM-面筋蛋白Tp分别增加3.49、3.18、2.8、2.64、2.59 ℃,在KGM取代度为0.5%时,KGM-面筋蛋白Tp增加的最多,表明KGM的添加使面筋蛋白的低热稳定性提高。ΔH表示面筋蛋白有序结构高低[30],由表1可知,当KGM取代度为2.5%时,KGM-面筋蛋白的ΔH增加量最高,为2.1 J/g。Wang Pei等[31]实验证明冻藏后面筋蛋白ΔH降低,面筋蛋白的有序结构减少,而添加KGM后面筋蛋白的ΔH增加,反映出KGM的添加使面筋蛋白结构变得更加有序,特别是在取代度为2.5%时,有序性相对较高。这主要由于KGM能够通过非共价相互作用和物理缠结改变面筋蛋白的构象,同时也能通过增强疏水相互作用促进蛋白质聚集,从而影响多肽链空间结构的重排[32]。
2.5 KGM对面筋蛋白游离巯基的影响

图4 KGM对面筋蛋白游离巯基含量的影响Fig. 4 Effect of KGM content on free SH content of gluten
作为面筋蛋白的功能组,游离巯基通常可通过形成二硫键参与蛋白质聚集行为。当面筋蛋白分子之间的二硫键断裂时,形成游离巯基,导致面筋蛋白的聚合物解聚[33]。从图4可以看出,与空白对照相比,在冷冻过程中加入不同水平的KGM后,面筋蛋白的游离巯基含量呈现出先降低后增加的趋势。当KGM取代度为0.5%时,面筋蛋白游离巯基含量从4.78 μmol/g降至4.55 μmol/g,这可能是由于KGM的自由基和亲核机制在链间二硫键诱导的面筋蛋白聚集中起重要作用[34]。当取代度从1.5%增加到4.5%时,面筋蛋白游离巯基含量分别增加0.13、0.58、0.51、0.61 μmol/g,这可能归因于活性羟基对KGM的D-葡萄糖和D-甘露糖的自由基清除作用,干扰了面筋蛋白二硫键的交换反应[32],从而对面筋蛋白中游离巯基含量产生影响。
2.6 KGM对面筋蛋白二级结构的影响

表2 冷冻期间添加KGM谷蛋白的二级结构占比变化Table 2 Change in secondary structure content of frozen gluten with added KGM%
蛋白质的二级结构主要通过非共价力维持,当环境发生变化时,蛋白质分子将重新排列,以达到最低能量,维持相对稳定的状态[31]。从表2可以看出,添加KGM会对面筋蛋白的二级结构产生影响。与对照组相比,KGM取代度为0.5%时,处理组的α-螺旋结构从28.08%下降到25.24%,降低了2.84%;当KGM取代度从1.5%增加到4.5%时,处理组的α-螺旋结构分别又增加了2.58%、4.21%、5.09%、7.45%,这可能是由于KGM的低取代度与面筋蛋白竞争性吸水而又无法充分相容造成面筋蛋白内部结构不稳定;β-折叠结构分别增加0.17%、3.25%、4.44%、5.06%、5.44%;β-转角结构分别下降了2.14%、3.98%、4.01%、5.28%、6.05%;无规卷曲结构也分别下降了5.17%、5.92%、7.71%、8.62%、8.59%。表明处理组改变了面筋蛋白二级结构的分布和形态,Linlaud等[24]研究也证实了KGM的添加使α-螺旋和β-折叠结构显著增多,有助于形成更紧凑的面筋蛋白结构。相较于β-转角和无规卷曲结构构象不稳定的特点而言,α-螺旋结构是一种稳定、强壮和有弹性的结构,有利于提高面团的弹性和硬度[35],β-折叠结构也是一种稳定的结构,所以KGM有利于改善冷冻面团的稳定性。这可能归因于KGM不仅可以作为亲水胶体修饰冰晶,使冰晶更小,分布更均匀,而且KGM可诱导多糖和面筋蛋白的—OH基团之间形成强烈的分子间氢键,产生更稳定的面筋网络结构[36]。
2.7 面筋蛋白微观结构分析


图5 面筋蛋白微观结构的扫描电子显微镜图(×500)Fig. 5 SEM images showing the microstructure of gluten added with different amounts of KGM (×500)
空白对照组(图5a)的网络结构密集,表面平坦,而所有样品处理组的面筋蛋白结构具有明显的海绵状结构和更大的孔隙,同时所有样品处理组的面筋蛋白微观结构显示出微米级的原纤维规律交联,第1层的原纤维形成连续的片状结构并形成了大量陷入网络中的空腔(图5b~f),与Zhou Yun等[37]的报道一致。随着KGM取代度的增加,面筋蛋白的结构更加蓬松、孔径更大。当KGM取代度为1.5%时面筋蛋白的孔径最大(图5c),这可能由于KGM掺入原纤维中,使其变平、伸长,有助于蛋白质链在其有序聚集发生之前展开[38],同时KGM不同于其他亲水胶体的结构,其能够自身形成糖网络,可以加固面筋蛋白网络结构,使其在冷冻环境下更加稳定。随着取代度的持续增加孔径开始回缩,这可能由于KGM具有很强的水合能力,使得过多的KGM能够与面筋蛋白竞争性吸水致使其结构开始回缩。
3 结 论
不同取代度的KGM对冷冻小麦面团面筋蛋白结构和功能特性存在一定的影响。KGM含有丰富的羟基,可束缚更多的水分子,提高了面筋蛋白的持水性。在KGM存在的情况下,面筋蛋白LMW-GS和醇溶蛋白α和γ型低分子质量区域的分子质量增大,并干扰面筋蛋白二硫键的交换反应,影响面筋蛋白的游离巯基含量。此外,KGM有利于提高面筋蛋白的黏弹特性和热特性,促使面筋蛋白中稳定性差的β-转角和无规卷曲结构转化为结构稳定的α-螺旋和β-折叠结构。面筋蛋白微观结构的变化也进一步证明了KGM对改善冷冻小麦面团面筋蛋白结构以及持水性、流变性、热特性功能特性的有效性。
