模拟叶片损伤条件下密胡杨幼苗的生理适应
2020-01-06师建银葛晓康白梦晗韩占江
孙 阳 师建银 马 楠 葛晓康 白梦晗 韩占江*
(1 塔里木大学生命科学学院/新疆生产建设兵团塔里木盆地生物资源保护利用重点实验室新疆阿拉尔843300)
(2 新疆生产建设兵团林业工作管理总站,新疆乌鲁木齐830000)
密胡杨(Populus talassica × P. euphratica)是以胡杨为父本、密叶杨为母本杂交选育获得的新树种[1],解决了胡杨杂交世界性难题[2,3,4]。密胡杨除具备胡杨耐寒、耐旱、耐盐碱、抗风沙的优良特性外,还具有无性繁殖能力强、生长速度快、材质好等优势[4,5]。在盐度为2.9%和pH值大于8.7的重盐碱条件下,密胡杨能够正常生长发育,具有外观整齐、生长速度较快、耐旱性强、景观效果好等特点[1],是干旱、盐碱条件下沙荒地区控制风沙的良好树种[6]。病虫害是林木上最常见的一种生物胁迫,是威胁林木生长的最大天敌,也是影响林业可持续发展的重要因素[7,8]。林木病虫害具有传染性,当某些地区出现病虫害时,它们会迅速蔓延到附近的植物,原有林木结构将被破坏,生物链被破坏,生态系统受到影响,极易造成巨大的经济损失[9]。近年来,在新疆南疆病虫害对树木的危害时有发生,部分团场防护林损失有增加趋势。受病虫害侵染后的林木会直接导致其体内活性氧和自由基的增加以及导致质膜损伤,抗氧化物质含量、保护酶活性均会发生变化,代谢失调,不仅影响植物的正常生长和发育,而且严重影响到它们的各种生理代谢活动。相应地,植物体内会通过逆境蛋白的表达、启动抗氧化防御系统、渗透调节等降低质膜系统的损伤。保护酶系统——超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、过氧化物酶(POD)等与渗透调节物质——可溶性糖、可溶性蛋白、游离脯氨酸等在植物的逆境适应中发挥重要作用,能提高细胞对逆境的抵抗能力[10,11]。鉴于此,通过分析保护酶活性、渗透调节物质含量及过氧化产物(MDA)含量变化,可以帮助了解植物在伤害条件下的生理调节机制。
作为新树种,密胡杨对伤害条件的适应能力关系到其推广面积和生态作用的发挥。基于此,采用人工模拟密胡杨叶片受伤害条件,通过分析损伤后密胡杨生理生化指标的变化,评估密胡杨对伤害的适宜能力,以期为科学管护密胡杨树苗,合理开发利用提供参考。
1 材料与方法
1.1 试验材料
试验材料为1年生密胡杨幼苗。
1.2 试验方法
1.2.1 密胡杨的前期处理及材料收集
试验于2018年生长季在塔里木大学园艺试验站进行,采用盆栽的方法。将长势一致的密胡杨幼苗分成3 组,每组10 株,1 组作为对照组,2 组作为伤害处理组,分别对密胡杨幼苗叶片进行20%和60%的模拟病虫害处理(均匀地将密胡杨幼苗上的叶片除去20%和60%)。伤害处理后,定期观察密胡杨幼苗的生长情况,保持三个月相同条件生长,然后取叶片,测定生理指标,3次重复。
1.2.2 生理指标的测定
相对电导率采用电导仪法测定,丙二醛(MDA)含量采用硫代巴比妥酸法测定,超氧物歧化酶(SOD)活性采用氮蓝四唑法测定,过氧化氢酶(CAT)活性采用高锰酸钾滴定法测定,过氧化物酶(POD)活性采用愈创木酚法测定,游离脯氨酸含量采用磺基水杨酸法测定,可溶性蛋白含量采用考马斯亮蓝G-250 法测定,可溶性糖含量采用蒽酮比色法测定[10]。
1.3 数据处理
利用Microsoft Excel 2010 软件进行数据分析和作图,应用DPS(9.01)统计软件进行方差分析及LSD检验(P<0.05)。
2 结果与分析
2.1 不同伤害条件下密胡杨幼苗相对电导率的变化
相对电导率是反映植物膜系统的一个直观数值,可以反映植物体内生物膜透性变化的情况,植物在遭受逆境时,细胞膜透性会增加致使电导率升高[10,11]。由图1 可以看出,与对照相比,在20%伤害条件下,密胡杨幼苗相对电导率无显著变化;但在60%伤害条件下,相对电导率显著增加,说明较高强度的伤害造成了密胡杨幼苗细胞膜的损伤。
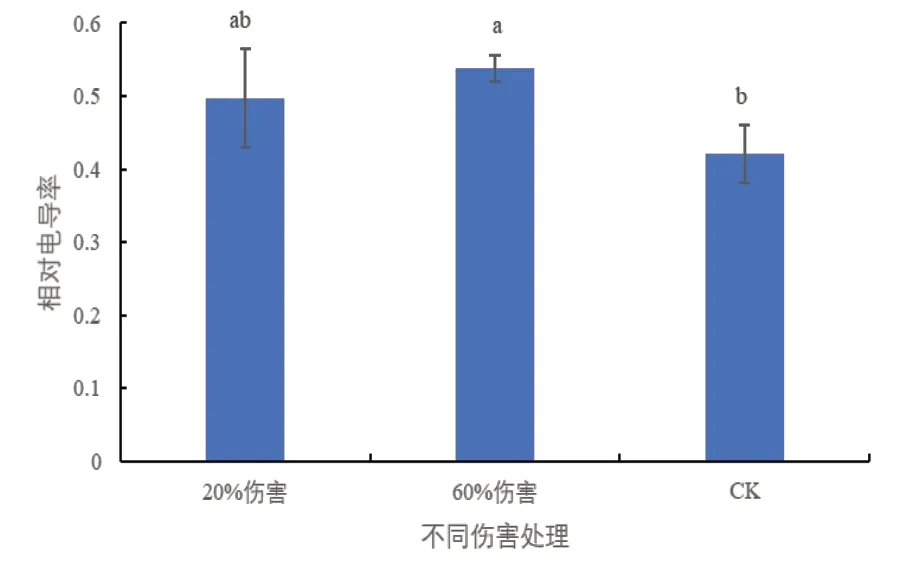
图1 不同伤害程度下的密胡杨幼苗的相对电导率
2.2 不同伤害条件下密胡杨幼苗MAD含量的变化
通常植物在条件不利时,生物膜脂会与自由基发生过氧化反应,MDA 是膜脂过氧化作用的产物之一,其含量高低与植物的抗性强弱成反比[10,11]。从图2 可以看出,与对照相比,20%伤害条件下,密胡杨幼苗的MDA含量无显著变化;在60%伤害条件下,密胡杨幼苗的MDA 含量显著增高,说明密胡杨幼苗的细胞膜脂发生了过氧化反应。
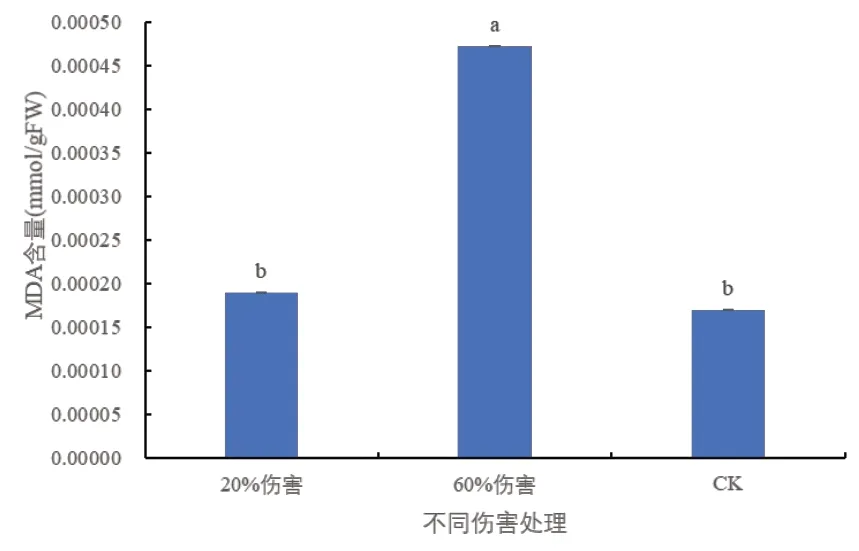
图2 不同伤害程度下密胡杨幼苗的MDA含量
2.3 不同伤害条件下密胡杨幼苗SOD活性的变化
SOD 在植物体内是一种可以抗氧化、抗衰老的酶类,对植物细胞具有保护作用[10,11]。由图3 可以看出,与对照相比,在20%和60%伤害条件下,密胡杨幼苗的SOD 的活性均未发生显著变化,说明在伤害条件下密胡杨幼苗的SOD活性保持稳定。
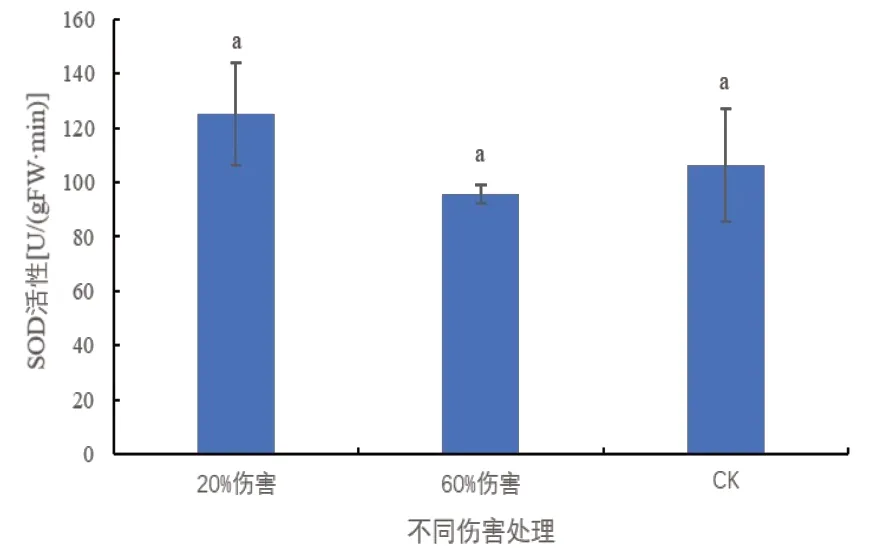
图3 不同伤害程度下密胡杨幼苗的SOD活性
2.4 不同伤害条件下密胡杨幼苗CAT活性的变化
CAT 可以清除体内活性氧,防止植物细胞在不良条件下的衰老等[10,11]。由图4 中可以看出,与对照相比,在20%伤害条件下,密胡杨幼苗的CAT 活性未发生显著变化;在60%伤害条件下,密胡杨幼苗CAT活性显著下降,说明密胡杨幼苗通过CAT 清除体内积累的活性氧的能力显著降低。
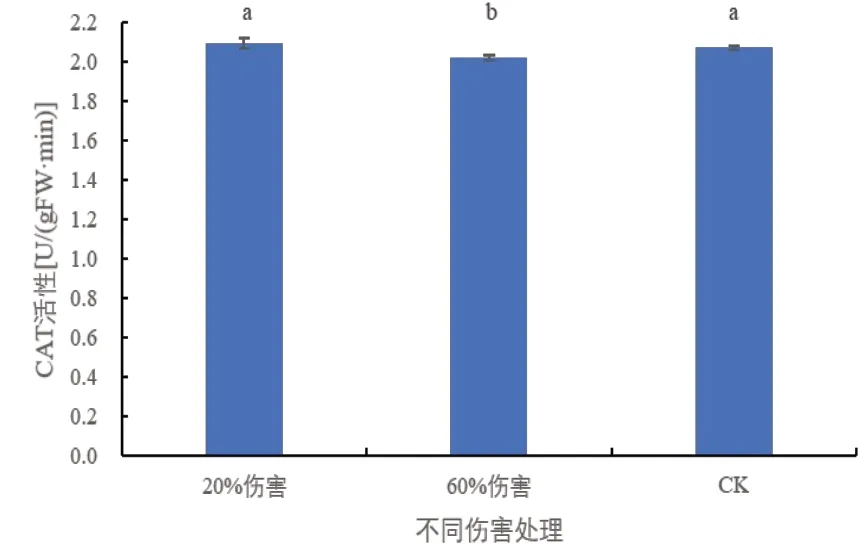
图4 不同伤害程度下密胡杨幼苗的CAT活性
2.5 不同伤害条件下密胡杨幼苗POD活性的变化
POD 是植物体内普遍存在的、活性较高的一种酶,它与呼吸作用、光合作用及生长素的氧化等均有密切关系[10,11]。由图5 中可以看出,与对照相比,在20%伤害条件下,密胡杨幼苗POD 的活性无显著变化;在60%伤害条件下,密胡杨幼苗POD 的活性显著升高,说明密胡杨幼苗通过提高POD活性来抵御伤害。
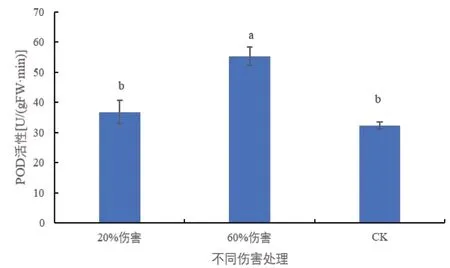
图5 不同伤害程度下密胡杨幼苗的POD活性
2. 6 不同伤害条件下密胡杨幼苗中游离脯氨酸含量的变化
脯氨酸是一种良好的渗透调节物质,通常以游离的状态存在,对植物进行渗透调节[10,11]。由图6 可以看出,与对照相比,在20%和60%伤害条件下,密胡杨幼苗的游离脯氨酸含量均未发生显著变化,说明脯氨酸合成对伤害条件不敏感。
2. 7 不同伤害条件下密胡杨幼苗可溶性蛋白质含量的变化

图6 不同伤害程度下密胡杨幼苗的游离脯氨酸含量
植物体内可溶性蛋白的含量通常与酶的活性相联系,可溶性蛋白含量的高低反映了植物体内的生理总代谢[10,11]。由图7可以看出,与对照相比,在20%伤害条件下,密胡杨幼苗的可溶性蛋白含量未发生显著变化;在60%伤害条件下,密胡杨幼苗的可溶性蛋白含量显著降低,说明密胡杨幼苗体内的蛋白代谢紊乱,分解大于合成。
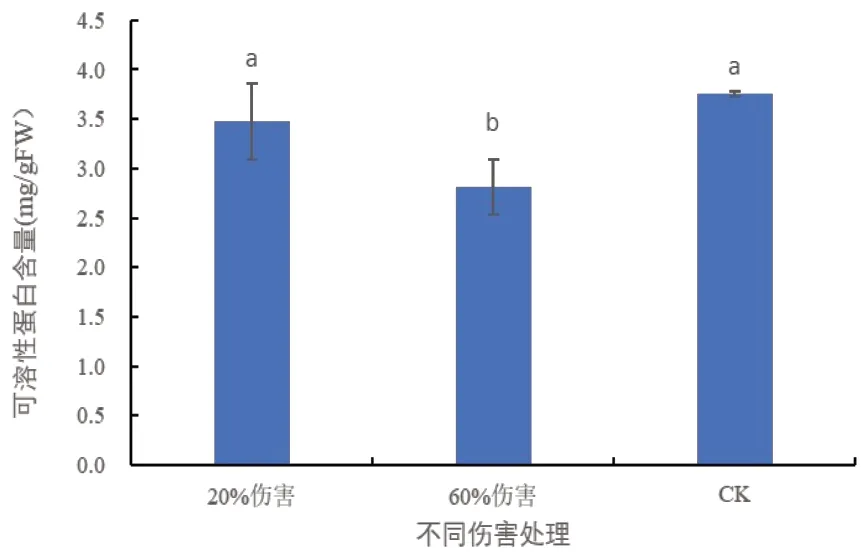
图7 不同伤害程度下密胡杨幼苗的可溶性蛋白含量
2.8 不同伤害条件下密胡杨幼苗可溶性糖含量的变化
可溶性糖可参与植物体内的渗透调节[10,11]。由图8 可以看出,与对照相比,在20%和60%伤害条件下,密胡杨幼苗的可溶性糖含量均显著降低,说明密胡杨幼苗的糖代谢紊乱,分解大于合成。
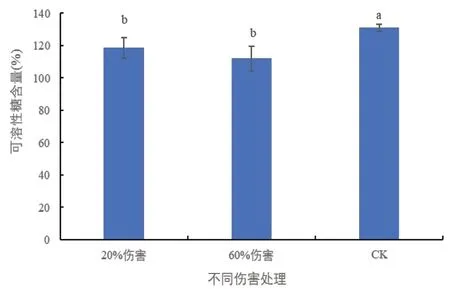
图8 不同伤害程度下密胡杨幼苗的可溶性糖含量
3 结论与讨论
在人工模拟自然生态对密胡杨幼苗伤害后(病虫害等),密胡杨幼苗的生理生化指标发生了一定变化。逆境会使细胞膜系统受到伤害,导致电解质大量外渗,因而相对电导率可反映细胞膜受损程度。MDA 是植物细胞内膜脂过氧化的产物,会引起细胞膜功能紊乱[12],MDA的积累量越少,植株受到的危害就越轻,抗性越好,反之,抗性越差。因此,相对电导率和MDA含量是常作为逆境胁迫下细胞质膜受到伤害程度的指标,通过分析其含量的高低,可以了解膜脂过氧化程度、膜系统受损程度以及植物抗性强弱[13]。本研究结果表明,在60%伤害条件下,相对电导率和MDA 含量均显著增加,说明较高强度的伤害会造成密胡杨幼苗细胞膜的损伤。
保护酶体系(SOD、POD、CAT)是植物体内的活性氧自由基清除机制之一,在逆境中能使植物通过提高其活性,来减少活性氧对植株的伤害,维持细胞膜的稳定[14]。其中,SOD 能够催化超氧化物的歧化反应,具有清除活性氧自由基的作用[15]。本研究中SOD 活性无显著变化,说明密胡杨幼苗对逆境抵抗能力与维持体内较高SOD 活性的能力有关[16];CAT能够调节活性氧伤害,但其调节能力是有限的,当胁迫达到一定程度后,其活性会降低,失去调节能力[17];POD 活性升高可以保护植物细胞膜的结构,其活性升高的原因可能是活性氧的增加刺激了保护酶系统的活性[18]。总体而言,在伤害条件下,密胡杨幼苗保护酶的活性调节机制比较复杂。
渗透调节的主要功能是维持细胞膨压和细胞膜稳定[11]。本研究发现,与对照相比,在20%伤害条件下密胡杨幼苗的可溶性糖含量显著降低;在60%伤害条件下,除脯氨酸含量升高,可溶性糖含量、可溶性蛋白含量均显著降低,这与已报道病虫侵染树木的变化趋势一致。其原因可能是植物受到伤害条件时产生了防御反应,而合成防御反应中的碳源主要由糖类提供[19]。由此可见,可溶性总糖含量的减少是植物在遭受生物逆境后,对代谢过程重新调整的结果[9]。可溶性蛋白同样作为渗透调节物质的一种,在逆境条件下,植物体内的蛋白质合成受到抑制。本研究发现,在20%伤害条件下,密胡杨幼苗会诱导合成新的蛋白,减弱伤害条件的不利影响;在60%伤害条件下,密胡杨幼苗可溶性蛋白含量显著降低,说明此时密胡杨幼苗的伤害已较为严重。游离脯氨酸是细胞质中重要的渗透剂和防脱水剂同时对植物体内的酶和膜结构有保护作用[20]。在正常生长条件下,植物体内的游离脯氨酸含量很低[21],但在逆境条件下脯氨酸含量会增加[22]。胁迫条件通常容易引起蛋白质分解,脯氨酸首先被大量地游离出来[23]。近年来的多项研究表明,脯氨酸的累积与植物的抗逆性间的关系在不同的植物中存在差异[24]。本研究发现,与对照相比,在20%和60%伤害条件下,密胡杨幼苗的游离脯氨酸含量均未发生显著变化,说明脯氨酸合成对伤害条件不敏感,在一定程度上反映了密胡杨对伤害条件的生理调控机制是由多种生理过程共同参与和决定的综合性表现。
综合以上指标的变化规律可以看出,密胡杨幼苗在20%伤害条件下尚能保持一定的自我调节、修复能力,而伤害程度达到60%时,其代谢发生紊乱,修复能力几乎丧失。因此,密胡杨对病虫害等伤害程度较轻时可以进行自身调节、修复,但在病虫害对其损伤较重的情况下则需要进行人工养护。
