海洋生物泵研究的若干新进展与展望
2019-12-19黄邦钦陈纪新
黄邦钦,邱 勇,陈纪新
(1.厦门大学近海海洋环境科学国家重点实验室、福建省海陆界面生态环境重点实验室、环境与生态学院,福建 厦门 361102; 2.泉州师范学院海洋与食品学院、福建省海洋藻类活性物质制备与功能开发重点实验室、近海资源生物技术福建省高校重点实验室,福建 泉州 362000)
海洋是地表系统最大的碳库,自工业革命以来海洋吸收了约48%的人为排放CO2,在全球碳循环中起着非常重要的作用[1-2]。海洋浮游植物占不到全球1%的光合自养生物生物量,但却贡献了近50%的初级生产力[3]。随着海洋碳循环研究的不断深入,相继提出了海洋“物理泵”(“溶解度泵”)、“化学泵”(“碳酸盐泵”)、“生物泵”和“微型生物碳泵”等储碳机制。海洋生物泵是指浮游植物光合作用将无机碳(CO2)合成颗粒有机碳(POC,即浮游植物细胞),通过自身沉降和浮游动物的摄食打包沉降等一系列复杂过程(包括初级生产、摄食、聚集、呼吸、矿化、沉降等)将碳从海表层输送出真光层或弱光层海的过程[4-5]。已有模型研究表明,如果海洋生物泵关闭,那么大气中的CO2浓度比率会增加0.02%,超过当前大气中浓度的40%,这将进一步加剧温室效应并加速气候变暖[6-7]。因此,海洋生物泵在全球碳循环与气候系统调控中起十分重要的作用。据估计,海洋生物泵每年去除超过100亿吨来自于海洋上层的碳,但其中只有约10%的碳输运到达深层[6]。已有研究表明,大约2/3海洋中碳的垂直梯度是由于生物泵而产生的,其余的1/3是由于溶解度泵产生的[7]。
较为系统地开展海洋生物泵研究可以追溯到20世纪80年代末启动的全球海洋通量联合研究(JGOFS),此后,多个全球大型海洋研究计划,如海洋生态系统动力学研究(GLOBEC)、海洋生物地球化学和海洋生态系统整合研究(IMBER)、海洋碳与生物地球化学(OCB)和海洋碳与气候变化(OCCC)等均涉及生物泵研究。海洋学家运用多种技术手段(如沉积物捕获器、同位素示踪等)获取了全球边缘海和大洋大量的实测数据,对生物泵生态过程和机制有了较好的认识,尤其是对真光层生物泵过程与调控机制方面取得了较好的进展。
然而,生物泵是个极其复杂的生态系统过程,影响生物泵效率的因素很多,比如初级生产过程、浮游生物(浮游植物和浮游动物)群落结构、浮游动物对浮游植物的摄食和微生物的矿化作用[7],此外,生物泵效率还取决于富含有机质的聚集体的形成和分解,以及POC聚集体与悬浮的“压载物”矿物质之间的相互作用[8]。通常生物泵效率较低, 而且时空变动很大,大部分在水柱上层通过光合作用固定的碳会在真光层(100~200 m)和弱光层(约200 m至1 000 m)的中上层被再矿化。一般来说,只有约5%~25%的净初级生产量输出真光层,在大洋荒漠区,通常小于10%净初级生产量输出真光层[9],但在极地海域可高达30%~100%[10]。通常仅有3%的净初级生产量可输出弱光层,抵达无光层(大于1 000 m)[8]。上层初级生产的碳约15%被细菌消耗[3],30%~70%被小型浮游动物摄食[11]以及20%~35%被中型浮游动物摄食[12]。此外,物理过程(如中尺度涡旋、上升流等)通过影响化学和生物群落特征而显著影响生物泵输出通量和效率[13-14]。在当今全球变化(如暖化、酸化、富营养化等)的背景下,生物泵效率也将受到影响,全球变化可能通过改变初级生产力、生物群落结构和微生物代谢等影响生物泵POC输出通量和效率。
1 浮游植物群落结构对生物泵的影响
浮游植物是全球海洋最主要的初级生产者,初级生产驱动了生物泵的运作。但初级生产力与生物泵POC输出通量并不是简单的线性关系,浮游植物群落结构、沉降特性以及通过食物链的传输等都会影响POC输出通量。浮游植物的沉降速率总体较低,通常只有小于1~10 m/d[15-17],显著低于其它较大颗粒的沉降速率[18-21]。由于体积较小且沉降速率很慢,沉降过程中容易被摄食和矿化,通常情况下浮游植物的直接沉降对POC的贡献并不高。直到最近20 a来的研究表明,在藻华发生的海区浮游植物可以直接快速沉降到海底,浮游植物的直接沉降输出才被认为是生物泵POC输出通量的主要组成部分[22]。此外,浮游植物群落的组成也会影响POC输出通量和效率,浮游植物通过影响沉降颗粒的压载物,进而也会影响POC输出通量和效率[23],如硅藻和颗石藻类可能对碳通量有很强的影响,因为它们的硅质外壳和颗石外壳提供了重要压载物,从而提高了颗粒物的沉降速率[23-24]。阿拉伯海和北大西洋的基于沉积物捕获器和234Th的生物泵研究结果表明,浮游植物群落中硅藻的比率与生物泵效率呈正相关关系[3, 10],Cai等(2015)在南海陆架和陆坡的研究亦有类似的结果,浮游植物中硅藻比例越高,POC输出通量越高,但海盆区则出现不一样的关系[25]。由于生物泵生态过程的复杂性(如群落结构演变与输出通量的时间不匹配,后者滞后)等原因,浮游植物群落结构与POC输出通量之间的上述关系规律的研究结果并不能用于推断其他地区的生物泵影响模式[10]。
浮游植物细胞的沉降速率受多种因素影响,包括各种形式的运动、细胞形态、生理状态和介质的粘度与密度,以及局部海水的动力学特性[26]。Bienfang等 (1984) 采用SETCOL管模拟沉降的方法,比较研究了29种不同浮游植物的沉降速率与细胞碳、氮含量和细胞体积的关系,结果显示浮游植物细胞沉降速率与细胞碳、氮含量和细胞体积3项因子均呈线性相关关系[27]。其中较小型浮游植物细胞(细胞直径小于20 μm)同种间沉降速率相近,沉降速率均较低,仅为0.15 m/d左右,而较大体积的浮游植物(细胞直径大于20 μm)不同种类的沉降速率变化范围是0.23~1.70 m/d,沉降速率与细胞体积呈线性正相关。在圣赫勒拿湾测量了在各种环境条件下不同组分的浮游植物组合的沉降速率,其中基于叶绿素a的浮游植物沉降速率介于0.00~0.91 m/d之间,而基于碳生物量的浮游植物沉降速率介于0.00~0.78 m/d之间,但是这二者的相关性很小[28]。浮游植物沉降速率与野外所测量的所有环境参数均无显著相关性,而与浮游植物种类组成显著相关,而后者受各种环境条件的制约。由于颗粒沉降动力学可能影响不同粒级浮游植物的生物量和生产力的垂直分布和物种演替模式。研究亦表明浮游植物的生理状态对其沉降速率有明显的影响[29]。通过分析不同浮游植物在不同环境条件下的沉降速率,可以进一步了解浮游植物群落结构的演变对生物泵输出通量和效率的影响[28]。不同生理状态(如生长期和衰老期)的浮游植物细胞沉降速率不同,其中活跃的浮游植物细胞沉降速率接近0~30 m/d,而衰老细胞的沉降速率比活跃期细胞快3~4倍[30]。研究还表明,光照[31]、细胞营养[15, 31]、细胞大小和细胞碳含量等均会影响沉降速率[30],进而影响生物泵POC输出通量与效率。
2 浮游动物群落结构对生物泵的影响
浮游动物摄食和粪球输出是海洋生物泵的关键过程。CO2进入海洋后,被光合浮游植物固定下来的POC在海洋生态系统中通常有两个归宿,一部分是初级生产者浮游植物直接被消费者浮游动物、游泳动物等摄食后进入更高营养的消费者体内,然后通过呼吸作用产生CO2在真光层循环。另一部分是浮游植物和浮游动物粪球颗粒直接向下沉降,在沉降过程中还可能与其他颗粒一起聚集(形成聚集体),然后以聚集体的形式向下沉降。在沉降过程中还会发生被再次摄食打包、降解导致再矿化等过程,最终部分颗粒物输出到深海中。浮游动物产生的粪球颗粒可显著促进POC的垂直通量,是生物泵的重要组成部分[32]。
浮游动物粪球颗粒对总POC输出通量的贡献变化很大(0%~ 99%),且受多种因素的影响[32]。浮游动物群落组成、垂直迁移行为和营养模式都可以影响粪球颗粒的丰度和组成,从而影响POC输出通量和效率[33]。Turner(2015)总结了全球浮游动物粪球输出通量研究结果,表明部分文献报道粪球输出通量贡献率很高,是POC输出通量的最重要来源,也有部分文献报道中粪球输出通量贡献率较低,还有一些报道中粪球输出通量对POC 的贡献具有可变性[34]。例如在赤道大西洋的400 m以上水层中,浮游动物粪球有机物质含量仅占总悬浮物质量浓度的4%,却贡献了400 m层的POC输出通量的99%左右[35]。在北大西洋5月春季藻华事件结束后,栖息于北大西洋深处的桡足类飞马哲水蚤(Calanusfinmarchicu)粪球颗粒占总POC沉降输出通量的92%[36]。在挪威附近海域200 m以上水层,粪球平均贡献了11%~37%,最高达66%的POC输出通量[37]。在加拿大的圣劳伦斯湾6—9月份浮游动物粪球颗粒POC输出通量占总POC输出通量的3%至接近100%,而在粪球POC输出通量最高时期占总POC输出通量的比率均高于66%[38]。此外其他一些研究表明,在棕囊藻(Phaeocystis)[39-40]和颗石藻(Coccolith)[41-42]的藻华期间磷虾(Euphausia)和桡足类动物粪球则是POC输出通量的主要贡献者。
浮游动物的摄食活动会影响颗粒物到达深海的速率,这种沉降的POC输出通量的大部分变化发生在海洋中“弱光层”内(即真光层至1 000 m)[43-44]。在海洋中层,浮游动物会消耗较小的沉降颗粒或悬浮颗粒,并将它们再打包成密度大、沉降速度较快的粪球颗粒[45]。粪球颗粒的输出也受季节性食物供应、摄食、食粪性行为和微生物再矿化等过程的影响[46-47]。在沉降过程中,粪球颗粒会通过被动物直接摄食碎裂成较慢或不容易沉降的颗粒或者被其它浮游动物重新吞食,还有些粪球颗粒与其他颗粒聚集在一起,形成海洋雪(简称海雪)[48]。粪球颗粒含有大量未消化或部分消化的物质,可供细菌和其它浮游微生物摄食并释放溶解有机碳(DOC),浮游生物和细菌改变颗粒特性并减缓碳的输出[43]。粪球颗粒的类型随着深度发生变化,可作为海洋中层浮游动物重新包装的指示[49]。
“海雪”是由浮游植物(包含植物碎屑)、海鞘类住囊虫丢弃的住屋、浮游动物粪球和其他杂质粘液等聚集而成的大于500 μm的肉眼可见的大型有机聚集体,是POC沉降输出的重要组成部分[17]。浮游植物向下沉降主要有两种路径,第一是被浮游动物摄食以后形成粪球,以粪球的形式沉降;第二是浮游植物直接沉降(例如硅藻、颗石藻类)或者与其他颗粒一起在一定条件下形成较大体积的聚集体,最终以聚集体的形式沉降输送到深海[34,50]。在藻华发生期间,浮游植物和其他颗粒一起易形成聚集体,这些絮状聚集体沉降速率的范围是从每天几十到几百米,极大地提高了浮游植物细胞的沉降速率和POC输出通量[51]。在东海近岸藻华区的研究发现,东海原甲藻(Prorocentrumdonghaiense)藻华期间,由于浮游植物大多以聚集体的形式集体沉降,东海原甲藻的平均沉降速率增加了10倍,而潜在输出通量增加了约100倍[22]。
浮游动物粪球颗粒对总POC输出通量的相对贡献还与所在海域的物理过程(如上升流)和浮游动物桡足类生命周期有关。研究认为在阿拉伯海中的浮游动物可以控制垂直通量的变化[52]。6月上升流开始时,上层大型桡足类大量捕食硅藻,防止硅藻大量繁殖,8—9月上升流季节结束时,最大垂直通量事件发生在桡足类迁移到深海中并进入滞育之后,此时未被摄食的硅藻成为沉积物捕获器中沉降颗粒的主要成分[53]。粪球颗粒对总POC输出通量的相对贡献比率可能会因厄尔尼诺而发生变化[54]。1997年1月,在1997—1998年洪保德海流北部厄尔尼诺之前,浮游动物粪球颗粒在300 m深的沉积物捕获器中占总POC的5%~10%,这是在浮游动物群落以桡足类为主的时期。在1997年7月,厄尔尼诺开始时,温跃层的加深导致上升流的营养盐输入减少,叶绿素水平降低。此时浮游动物中有丰富的纽鳃樽和磷虾类, 来自磷虾的浮游动物粪球占总POC输出通量的24%~38%。1998年1月,在厄尔尼诺后期,桡足类再次占优势,浮游动物粪球POC输出通量对总POC输出通量贡献下降至2.7%~11.3%[54]。
3 全球变化对海洋生物泵的影响
据推测,生物泵最早在海洋中运作可追溯至5亿年前的显生宙[55-56],因此,生物泵与气候变化的相互作用(影响、响应和反馈)已经历了漫长的地球历史进程。在未来全球变化背景下,海洋暖化、酸化和富营养化条件下生物泵将如何运作是我们必须揭示的重要课题,但目前我们对此了解还很少(图1)。
海水变暖通常增强水体层化,减少深层营养盐的向上输运,上层营养盐浓度降低,初级生产力降低,生物群落结构发生改变,由大粒级的硅藻变为小粒级的聚球藻(Synechococcus)和原绿球藻(Prochlorococcus),进而使生物泵输出通量减少。但海水暖化亦有增加生物泵输出通量的案例,如全球变暖提高风强度或风暴(如台风)几率增加,提高了高营养盐低叶绿素(HNLC)区的铁浓度,并增加透明胞外多聚物(TEP)的生产和硅藻的聚集,生物泵输出通量增加[34]。
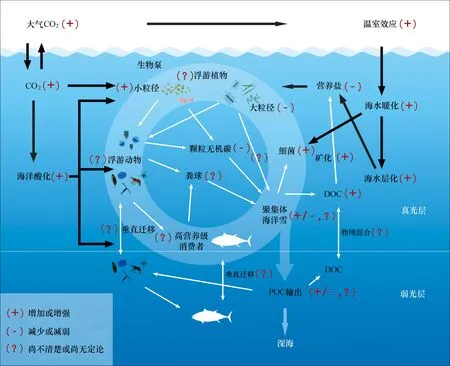
图1 海洋生物泵及其对全球变化的响应Fig.1 Biological pump and its response to the global change
海水变暖加快了生态系统的生物代谢过程(如异养微生物的分解速率),这将导致POC输出通量的降低[57-58]。变暖也改变上层海洋生态系统,从以硅藻-中、大型浮游动物为优势的系统转变为以微微型浮游植物-小型浮游动物为优势的系统,进而减低POC输出通量。模型的研究亦显示,在未来持续暖化背景下,全球海洋初级生产力和POC输出通量将降低[59-60]。海水暖化也促进海雪的形成和有机物的聚集。研究发现温度升高使硅藻聚集加快,类似TEP在较高温度下浓度升高的特性[61],然而升温也刺激细菌的生长和代谢,导致聚集物的快速降解。因此,暖化不仅提高了聚集体和海雪的形成速率,而且也促进了颗粒物的降解速率,这导致相应的生物泵的增强或减弱。
随着大气中CO2含量上升,pH下降,海水酸化。海水中CO2浓度升高对生物泵的影响亦很复杂,虽然CO2浓度升高能促进CO2限制的浮游植物光合作用,但也改变了浮游植物群落组成和总初级生产力,同时海水酸化使得碳酸钙等压载物容易被降解而减少输出,因而生物泵效率是提高还是降低,目前仍然没有确切的结论[34]。
海洋酸化将降低钙化浮游生物(如颗石藻、有孔虫等)的钙化强度,这是颗粒有机碳的压载物,如此将导致POC输出通量和生物泵效率的降低[62-66]。酸化亦改变主要营养元素的循环,特别是氮,但目前已有的结果相互矛盾。有结果表明,酸化促进了蓝藻的固氮作用、提高了反硝化作用、抑制了消化作用[67-68]。但新近的研究结果表明,通过区分了海水酸化的CO2效应和pH效应,发现CO2升高对束毛藻(Trichodesmium)的促进作用小于海水pH下降对其的抑制作用,导致海洋酸化的净效应为抑制束毛藻的固氮和生长,南海天然固氮生物群落的研究亦呈现一致的结论[69]。通过对海水培养基成分的分析并结合系统的受控培养实验,发现先前报道的海洋酸化对束毛藻固氮的促进作用,很可能是由于相关研究使用的人工海水培养基受铜和氨的污染所导致的假象——酸化降低了过量的铜和氨对束毛藻的毒性作用,使得其生长和固氮速率加快[70]。
细菌矿化对生物泵POC输出有重要影响,但目前高CO2或酸化对此影响的研究较少,在富营养化生态系统,围隔实验结果表明,在高CO2处理组,附着态和浮游态细菌的呼吸速率在平台期显著下调(约31%),而细菌生产力无显著变化,表明在CO2加富条件下,细菌碳同化的效率发生改变,总需碳量下调,更多吸收的碳用于自身生长而用于呼吸消耗的比例减少。自养和异养类群代谢速率的同时变化,促进整个生物群落净生产力升高,说明CO2加富效应将会对近岸生态系统碳循环产生深远的影响[71]。
未来的海洋暖化和酸化要同时改变海水、碳酸盐、层化、混合、营养盐和生物群落结构,但这种变化在全球不同区域而不同[7,57,72]。在这种多因素(包括温度、光照、营养盐、CO2、酸化、生物群落结构等)复合影响下,要想从各自的因子来预测生物泵的变化是很困难的[67-68,73-74]。因此,海洋学家要预测下个世纪生物泵的准确变化(变强或变弱)是很难的。受多种相互作用参数和协同效应的影响,这些参数都会影响生物泵的输出通量和储存量。
海水暖化和富营养化将改变浮游植物的群落结构,进而可能影响生物泵效率。Xiao等(2018)基于东海14 a(2002—2015年)累积的23个现场航次的大量浮游植物光合色素数据,采用多种统计学手段,建立了东海硅藻和甲藻对主要环境因子的响应模式。模型显示,硅藻和甲藻对温度和营养盐(氮、磷及其比值)变化的响应模式不同:硅藻偏好低温和高营养盐,而甲藻对温度和营养盐相对不敏感,但倾向于低磷和高氮磷比的环境。在假设温度和氮磷比(取对数)各升高两个单位的情境下,模型预测东海约60%的区域将出现硅藻生物量的下降,约70%的区域甲藻生物量将升高;变化最大的近岸区域硅藻生物量将降低19%,甲藻生物量将升高60%[75]。
量化生物泵的效率是构建全球碳循环模型的先决条件[76]。然而,在最近的海洋变暖和酸化对生物泵的预期影响模型中,关于上层生态系统如何应对这种变化仍存在很大的不确定性[77-78]。最近的一些模拟表明,过去10 a,全球初级生产力和生物泵碳输出已经呈下降趋势[79-84]。近几十年来,海洋变暖导致大气中CO2含量增加,从而加剧了大气和海洋变暖。
4 研究展望
针对当前海洋生物泵研究的概况及其未来全球变化情景下的生物泵变化趋势,本研究认为应该加强如下几个问题的研究。
(1)边缘海不同生态系统的生物泵结构、输出通量和影响机制。通常认为驱动海洋生物泵的决定性因素是浮游植物初级生产力,然而近年来很多研究发现两者并不存在简单的线性耦合关系。海洋上层食物网结构,特别是浮游动物的摄食打包、垂直迁移、细菌生产和呼吸等各种生物类群的生命活动都被认为是引起这种非耦合关系的因素。相对于大洋生态系统的生物泵研究,目前对边缘海的研究还十分肤浅。通过对边缘海不同生态系统(如陆架、陆坡和海盆等)生物泵结构的比较研究,阐明其调控机制。
(2)海洋全水柱生物泵研究。过去30 a来,海洋生物泵研究主要集中在真光层,最近的研究指出有机碳在弱光层的传递是个极为复杂的过程,主导这个过程的是以细菌和浮游动物等为主的中层生物群落,这些生物对有机碳的消耗、转换、矿化和再生产过程,极大地影响了中层水的碳循环过程和POC输出。然而,这样的研究还非常缺乏。
(3)全球变化情景下的生物泵变化趋势研究。当前以变暖为主要特征的全球变化在持续加剧,而且大气中CO2持续升高,pH下降导致海洋酸化,近岸富营养化和低氧(缺氧或无氧)已在进一步加剧。上述全球变化已经并将继续导致生态系统结构和功能发生明显转变,进而可能对生物泵储碳产生深远的影响,但对其影响机理和效应还不清楚,需要加强研究[34]。
(4)采用一些新的研究方法与技术。传统的方法(234Th-238U、漂浮式沉积物捕获器)比较适合真光层POC输出研究,而针对弱光层的研究需要探索新方法和技术的研究与应用,如210Po-210Pb[85-86]和中性浮力沉积物捕获器[87],可探讨采用漂浮式-中性浮力-锚链式沉积物捕获器(分别适合真光层-弱光层-深水层)组合集成模式开展全水柱生物泵POC输出通量研究。此外水下图像剖面仪(Underwater Vision Profiler, UVP)可应用于颗粒测定和估算沉降的研究[88-89]。
(5)构建更完善的生物泵模型并提高预测精度。目前大多生物泵模型只考虑由于重力作用导致的POC沉降过程,忽略了包括浮游动物垂直迁移、物理过程(如垂直混合、中尺度涡等)对POC以及DOC对生物泵的贡献[90]。同时目前预测结果也存在很大的不确定性。例如,对于真光层界面,目前几种模型预测的全球输出生产力的不确定性范围为5~12 pg/a(以C计),这个与每年由于人类活动新增进入全球海洋的CO2量相当。在今后的模型研究中,一个是构建更加完善的生物泵输出的机理模型[91],尤其是阐明中深层浮游动物垂直迁移过程贡献[92],并结合全球环流模型,全面评估不同物理和生物过程对生物泵的贡献。另一途径是基于观测数据的逆向模型,即利用大规模的现场实测数据,通过统计学或借助机器学习的手段(如人工神经网络、深度学习等)来训练算法[93],模拟复杂的生物泵过程,提高模型预测度来更加精确定量生物泵对全球气候变化的贡献。
