几种环境因子对崖州硬皮豆种子萌发的影响
2019-12-19虞道耿陈银华
曹 敏,张 瑞,钟 婷,虞道耿,陈银华,骆 凯
(1. 海南大学热带作物学院,海南 海口 570228;2. 中国热带农业科学院热带作
乡土草的驯化选育和推广利用不仅是解决我国牧草新品种紧缺的快捷和可行之途,还是一项符合国际发展趋势的长期性工作[1]。摸清不同乡土草种生物学和生态学特性,尤其是种子萌发和幼苗生长特性,是其驯化选育、高效栽培和推广建植的重要前提。因为种子萌发和幼苗生长时期是植物生活史中最敏感和脆弱的阶段,易受多种环境因子的综合影响[2-3];加之种子萌发特性在物种间具有很大的变异,共同决定了植物进入生态系统的时间和幼苗期耐受生存环境的能力[4]。因此,研究多环境因素对种子萌发的影响,有助于掌握和预测物种的时空分布、建植率和收获量等信息,也为乡土草优质栽培和新品种推广提供有用的参考数据[5-6]。
硬皮豆(Macrotyloma uniflorum),豆科硬皮豆属一年生乡土草,具抗旱、耐贫瘠、抗病虫害、生长势强、粗放管理等优点,被认为是一种优良的绿肥和饲料作物[7]。有报道称,该物种的干种子蛋白质含量为23%~26%,脂肪含量0.9%左右,也是海南本地的一种低脂、高蛋白食用豆类食品[8]。崖州硬皮豆出苗快、长势强、叶量多,与田间杂草竞争力强,可作为种木薯(Manihot esculenta)或其他果园优良的行间控草作物,应用前景广阔。
关于崖州硬皮豆,目前仅见生物学特征和营养栽培研究的报道。崖州硬皮豆为海南特有的农家品种,具有多项优良性状,其叶片具有敏感的感光性膨压运动特性,能够调节叶面水分蒸腾强度,根系发达,根瘤丰富,发展崖州硬皮豆生产可对海南西南部秋冬旱季土地起到用养结合的良好效果;同时还可以作为常规和基因工程育种的宝贵材料[7,9]。崖州硬皮豆的驯化选育是乡土草资源开发利用的一项科研成果,但相关的生物学和生态学特性掌握甚少,这将成为全面发掘该品种生产和生态服务功能的阻碍。基于此,本研究以采自海南岛的崖州硬皮豆为材料,在实验室模拟条件下分析不同温度、光照、水势、盐分和埋深对种子萌发和出苗的影响,探讨种子萌发特性对环境变化的适应对策,为崖州硬皮豆种子学研究奠定基础,也为引种驯化和开发利用提供依据。
1 材料与方法
1.1 试验材料
崖州硬皮豆种子由中国热带农业科学院提供。种子于2019 年1 月采于海南省东方市感城镇(18°50′ N,108°38′ E),经人工清选干燥后,置于中国热带农业科学院热带牧草研究中心种子库(4 ℃)贮存备用。
1.2 试验方法
1.2.1 种子千粒重测定
随机数取100 粒种子称其重量,重复8 次,计算平均值。最终求得崖州硬皮豆的千粒重为(27.5 ±0.3) g。
1.2.2 种子硬实率测定
采用纸上萌发法测定所有供试种种子硬实率,随机数取50 粒饱满种子置于垫有双层滤纸、直径为11 cm 的干净培养皿中,4 次重复,将培养皿置于25 ℃ (光照12 h,黑暗12 h)条件下培养,适时补充水分以保持萌发床湿润,第28 天统计未吸胀种子数(种子吸水3 d 后,所有吸水种子数目均不再增加)。最终求得崖州硬皮豆的硬实率为(32.0 ±3.6)%。后续试验在开始前,种子均用解剖刀人工切破种皮,以破除硬实,之后将破除硬实的种子贮藏在室温条件下,一周内使用。
1.2.3 种子萌发的温度和光照设置
同上使用纸上萌发法,分别置于恒温15、20、25、30、35、40、45 ℃和变温10 ℃/25 ℃、15 ℃/30 ℃、20 ℃/35 ℃条件下进行培养。各温度处理下分别设置光暗交替(12 h/12 h)和黑暗(24 h)两个处理,其中黑暗处理采用双层锡纸包裹培养皿模拟全黑暗条件的方法,每个处理设置4 个重复。以胚根突破种皮作为判断种子萌发的标准,为得到更详细的种子萌发动态,在种子吸水后萌发速率最快的时间段(前48 h)增加统计次数,之后每24 h统计一次萌发数,试验持续两周。
1.2.4 水势胁迫和NaCl 胁迫设置
同上使用纸上萌发法,在30 ℃、12 h/12 h 光暗交替的培养条件下,根据Michel 和Kaufmann[10]的方法及计算公式分别设置水势为0 (CK)、 -0.2、-0.4、 -0.6、 -0.8、 -1.0、 -1.2 MPa 的PEG-6000溶液。相同培养条件下,分别设置0 (CK)、50、100、150、200、250、300、400 mmol·L-1的NaCl溶液。每个处理设置4 个重复,每重复50 粒种子,每24 h 统计一次,持续两周,萌发标准同上。试验期间,每隔2 d 更换一次溶液以保持相对恒定的溶液浓度。
1.2.5 埋深设置
采用盆钵法,分别设置0、1、2、3、4、5、6、7 cm 共8 个埋深梯度,每个处理设4 个重复,每重复20 粒种子。试验基质为农田表层土壤,经130 ℃高 温 消 毒 处 理6 h。每24 h 统 计 一 次 出 苗率,以崖州硬皮豆子叶突出表层视为出苗,连续5 d没有种子出苗视为试验结束,试验周期为21 d。
1.3 数据分析
1.3.1 萌发和出苗指标
萌发(出苗)率 = (n/N) × 100%;
式中:n 为全部萌发(出苗)的种子数,N 为供试种子数。
萌发速率:采用Germinator 软件包计算萌发达到非硬实种子数50%所需的时间(t50),种子萌发速率用1/t50表示。
1.3.2 种子萌发的温度模型
采用种子萌发速率与温度(T,℃)模型,计算公式[11-12]为1/t50= k + mt。根据拟合出的方程式计算出最低临界温度(Tb)、最适萌发温度(To)以及最高临界温度(Tc)。
1.3.3 统计分析与作图
采用SPSS 20.0 进行统计分析,根据试验设计进行独立样本T 检验或双因素方差分析(ANOVA),采用Duncan’s 法对平均值进行多重比较。所有数据的录入和作图均使用Excel 2010 软件。
2 结果
2.1 温度和光照互作对崖州硬皮豆种子萌发的影响
温度、光照及二者互作对崖州硬皮豆种子萌发速率有一定影响,其中温度对种子萌发速率(1/t50)影响极显著(P<0.01),而光照对萌发速率(1/t50)的 影 响 不 显 著(P > 0.05) (表1)。恒 温 条 件下,崖州硬皮豆种子萌发率随温度升高,整体呈先上升后下降的趋势。萌发温度在低于20 ℃和高于40 ℃时显著下降(P<0.05),在10 ℃和45 ℃条件下种子均不萌发。萌发率在10 ℃/25 ℃、15 ℃/30 ℃和20 ℃/35 ℃之间差异不显著(P > 0.05),3 个变温均显著低于20、25、30 和35 ℃恒温(P<0.05),但显著高于低温10 、15 ℃和高温45 ℃ (P<0.05)。仅在15 ℃时,光照条件下的崖州硬皮豆种子萌发率显著高于黑暗条件下的(P<0.05),而在其他温度条件下,光照和黑暗对种子萌发率的影响均不显著(P > 0.05) (图1)。
光暗交替(12 h/12 h)和黑暗(24 h)下,崖州硬皮豆种子萌发速率与恒温温度的线性关系表现出相似的结果。随着温度的升高,种子萌发速率(1/t50)呈现明显的先升高后降低的趋势;种子萌发速率(1/t50)在0 ~ 30 ℃区间里与温度呈直线正相关关系,在30 ~ 45 ℃时与温度呈直线负相关关系(图2)。根据种子萌发速率(1/t50)和温度之间的线性关系方程,推算出光照条件下Tb、To、Tc分别为11.0、32.8 和45.5 ℃,黑暗条件下Tb、To、Tc分别为13.4、32.4 和45.9 ℃。
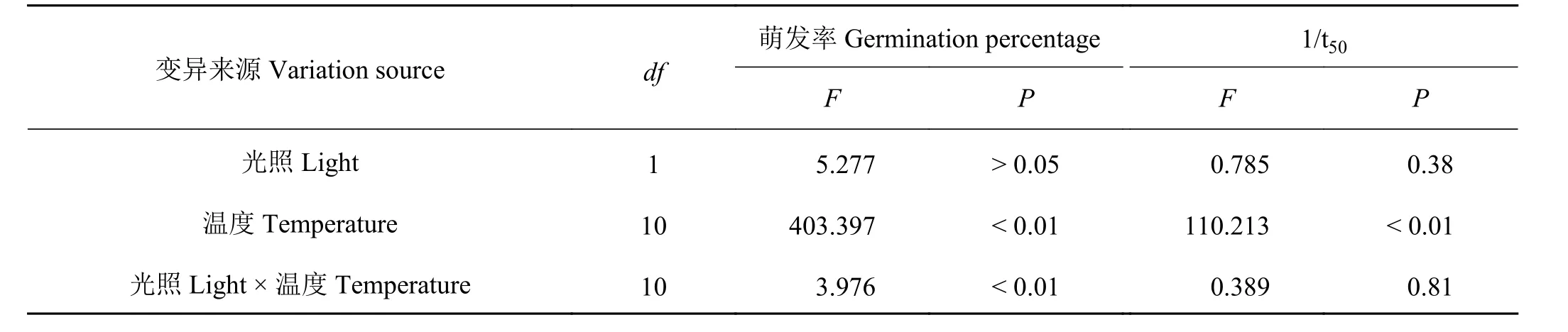
表 1 温度、光照及其互作对崖州硬皮豆种子萌发影响的双因素方差分析Table 1 Two-way ANOVA for the effects of temperature, light, and their interaction on the germination of Macrotyloma uniflorum ‘Yazhou’

图 1 光照和温度对崖州硬皮豆种子萌发率的影响Figure 1 Effects of light and temperature on the germination percentage and index of M. uniflorum ‘Yazhou’
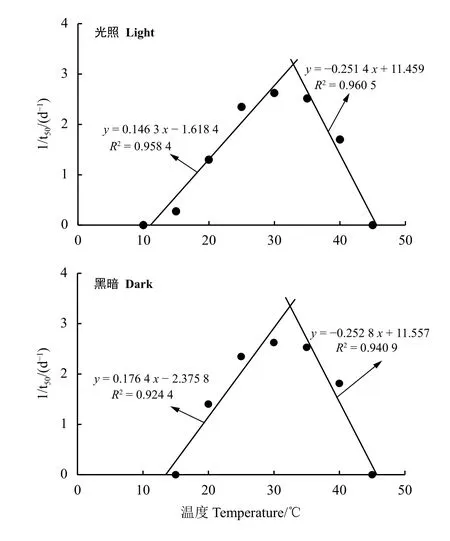
图 2 种子萌发速率(1/t50)与萌发温度的线性关系Figure 2 Linear regression of germination rate (1/t50)as a function of temperature
2.2 水分胁迫和NaCl 胁迫对崖州硬皮豆种子萌发的影响
崖州硬皮豆种子萌发率和萌发速率随渗透势胁迫的加剧而降低(表2)。水势在-0.4~0 MPa 的种子萌发率无显著差异(P > 0.05),且均超过99%。水势从 -0.6 MPa 开始,种子萌发率显著降低(P <0.05),达到 -1.2 MPa 时,萌发率仅为6.5%。
随着NaCl 浓度的增加,崖州硬皮豆种子的萌发率呈逐渐下降的趋势(表3)。当NaCl 溶液浓度低于200 mmol·L-1时,崖州硬皮豆种子的萌发率相比较未处理(CK)条件下无显著变化(P > 0.05);而NaCl 溶液浓度处于250~400 mmol·L-1时,崖州硬皮豆的萌发率被显著抑制 (P<0.05)。崖州硬皮豆种子萌发速率(1/t50)与NaCl 浓度呈明显的负相关关系。
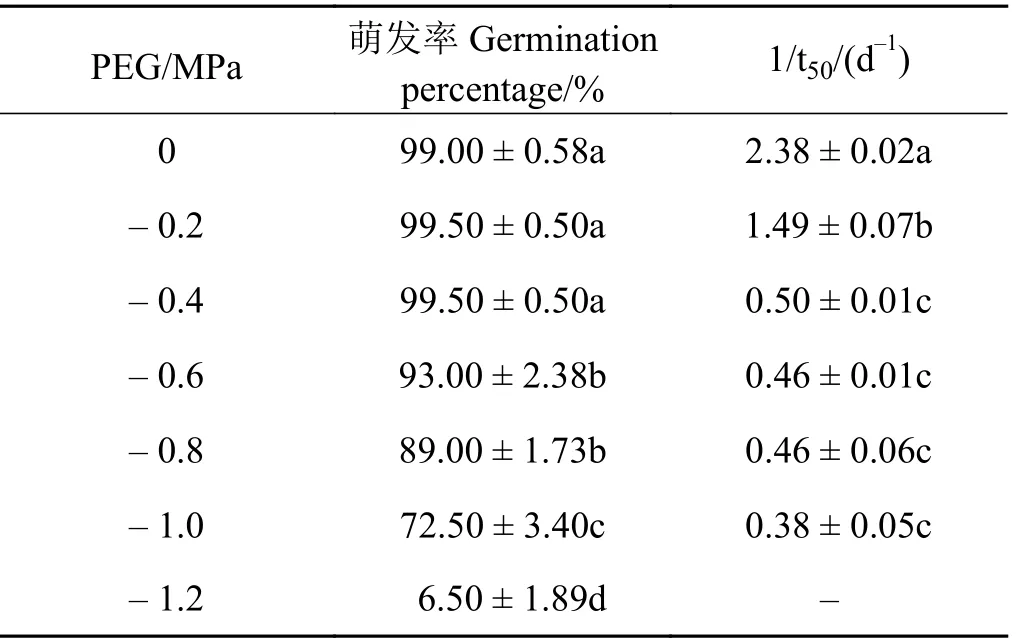
表 2 水分胁迫对崖州硬皮豆种子萌发率和1/t50 的影响Table 2 Effects of water stress on germination percentage and 1/t50 of M. uniflorum ‘Yazhou’
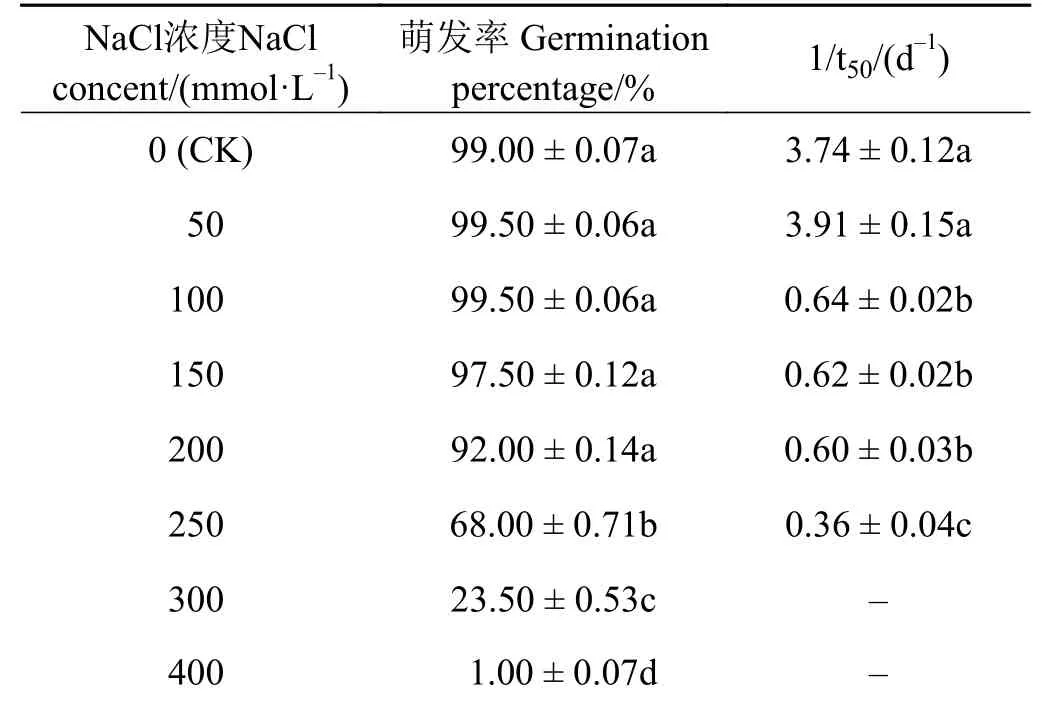
表 3 NaCl 胁迫对崖州硬皮豆种子萌发率和1/t50 的影响Table 3 Effects of NaCl stress on germination percentage and 1/t50 of M. uniflorum ‘Yazhou’
崖州硬皮豆种子萌发率和萌发速率随渗透势胁迫的加剧而降低(表2)。水势在-0.4~0 MPa 的种子萌发率无显著差异(P > 0.05),且均超过99%。水势从 -0.6 MPa 开始,种子萌发率显著降低(P <0.05),达到 -1.2 MPa 时,萌发率仅为6.5%。
2.3 埋藏深度对崖州硬皮豆种子出苗的影响
随着埋藏深度的增加,崖州硬皮豆种子出苗率呈现先上升后下降的趋势(图3)。埋藏深度在0~6 cm 时,崖州硬皮豆均有不同程度的出苗率,在1 和2 cm 埋深条件下出苗率较高,之后随着埋藏深度的增加显著下降(P<0.05),达到7 cm 时种子出苗率为0。埋藏深度在0 cm 时的出苗率显著低于1 cm 的(P<0.05)。
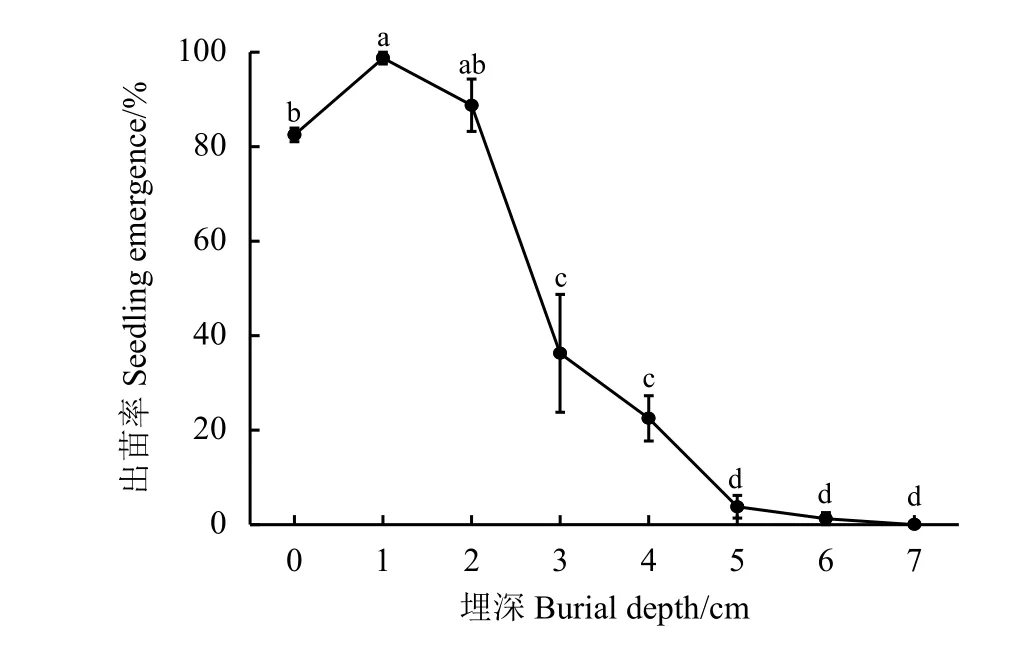
图 3 埋藏深度对崖州硬皮豆种子出苗率的影响Figure 3 Effects of burial depth on seedling percentage of M. uniflorum ‘Yazhou’
3 讨论
3.1 温度和光照对崖州硬皮豆种子萌发的影响
种子萌发是一个极其活跃的生理生化过程,伴随着一系列酶催化的物质代谢反应,而温度影响酶的活性,在最适温度范围内,酶的活性最强,因此适宜的温度成为影响种子萌发的主要因素之一[13]。这一范围内,温度过低或过高都不利于种子的萌发,通常使用温度三基点(最低、最适、最高)来描述一个物种所能萌发的温度范围[14]。本研究根据种子萌发速率与萌发温度的线性回归方程推算,在11~46 ℃内崖州硬皮豆可萌发,且在20~ 40 ℃下具有91%以上的萌发率,可见崖州硬皮豆种子具有广阔的萌发温幅,能在较大的温度范围内建植。多数情况下,变温环境相比较恒温环境更有利于种子萌发。以羊草(Leymus chinensis)种子萌发特性最为典型,在28 ℃或35 ℃恒温条件下,种子几乎不发芽,当处于5 ℃/28 ℃、5 ℃/35 ℃和16 ℃/28 ℃变温时,种子萌发率可超过80%[15]。但是,变温处理对崖州硬皮豆种子萌发不仅没有影响,甚至还起到抑制作用,这可能是避免种子在温度波动较大的地表萌发的一种适应机制,与长芒草(Stipa bungeana)[16-17]的研究结果相似。在亚适温条件下,相比较15 ℃恒温,崖州硬皮豆的萌发率在10 ℃/25 ℃和15 ℃/30 ℃变温时更高,说明变温减缓了低温对种子萌发的抑制作用。
不同植物种子萌发对光照的需求在种间存在较大差异,以形成对早期生境的生态适应性,保证尽可能多的幼苗存活与建成[18]。一般认为,种子萌发对光照的响应主要由遗传特性决定,但实际情况往往更为复杂,比如温度可能在一定程度上改变种子萌发的光需求[19]。多数种子萌发的光控试验是在适宜的温度、水分和O2条件下进行的,但自然界各生境特征差异大且易出现极端环境,研究光照和温度、水分、O2等环境因子的耦合作用对种子萌发影响,对光调控种子萌发机理的深入了解具有重要的意义。本研究表明,光照对崖州硬皮豆种子萌发的影响因温度不同而异。在适宜的萌发温度范围内,崖州硬皮豆种子萌发率和萌发指数在光照和黑暗下差异不显著(P > 0.05),说明崖州硬皮豆属于光中性种子。Metcalfa 和Grubb[20]对来自东南亚低地雨林的140 种植物种子进行研究,发现种子萌发的需光性随种子大小的增加逐渐减弱,崖州硬皮豆对光照需求小的特性可能与其种子较大(千粒重27.5 g)有关。在15 ℃的低温时,光照却显著促进崖州硬皮豆种子萌发(P <0.05),说明低温诱导崖州硬皮豆形成强迫性休眠,而光照可能作为打破强迫性休眠的信号因子[5,21-22]。
3.2 水分和盐胁迫对崖州硬皮豆种子萌发的影响
水分胁迫使外界环境形成大于种子内部的渗透势,这种差异将导致种子吸水速率减缓、萌发抑制或发芽延迟[23]。种子萌发对水分胁迫的反应与植物抗旱性相关,能在一定程度上反映该物种在干旱环境下的生存能力。本研究发现,崖州硬皮豆在PEG 6000 胁迫水势达到 -0.4 MPa 时,种子萌发率与对照相比未发生改变,这与Hu 等[16]在窄叶野豌豆(Vicia angustifolia)和‘兰箭3 号’箭筈豌豆(V.sativa‘Lanjian 3’)上的研究结果相似。轻度干旱胁迫下,植物种子会通过改变可溶性糖和脯氨酸等调节物质以维持相对正常的细胞膨压,保证萌发生理过程顺利进行[24]。崖州硬皮豆萌发速率(1/t50)在 -0.2 MPa 时就已显著下降(P<0.05),说明萌发速率对水分胁迫的变化与萌发率相比更为敏感,其原因可能是通过降低萌发速率,避免种子在水分缺乏的环境中大量萌发[3]。在剧变的环境中,这一机制可以提高种子较大且萌发吸水量较高的崖州硬皮豆幼苗的成活率。在盐胁迫中同样得到相似的结果。
盐分是土壤的重要组成部分,过量盐分会抑制植物生长和发育,进而导致作物产量和质量的下降[25]。盐分对种子萌发阶段的影响程度决定该物种适应广泛分布的盐渍化土地的能力,因而备受关注。景艳霞和袁庆华[26]发现,紫花苜蓿(Medicago sativa)种子在150 mmol·L-1NaCl 浓度条件下的萌发率显著低于对照(P<0.05),当浓度达到300 mmol·L-1时,种子萌发率为0。卢艳敏等[27]认为,白三叶(Trifolium repens)对NaCl 的耐受浓度阈值小于300 mmol·L-1。汪永平等[28]对5 份草木樨(Melilotus)种质进行耐盐性评价,结果表明,NaCl 浓度达到200 mmol·L-1时所有种质的萌发率相比较对照均显著(P<0.05)降低。本研究发现,当盐胁迫在200 mmol·L-1的NaCl 浓 度 以 下,对 崖 州 硬 皮 豆 萌发率没有显著的抑制作用(P > 0.05),与上述报道比较,崖州硬皮豆种子表现出较强的萌发耐盐性,具有在盐渍化地区推广的潜质。适应生态理论认为,植物种子萌发特性主要形成于原生境的适应过程中[29],崖州硬皮豆种子萌发对盐分限制因子的响应特征可能是长期适应海滨盐渍化区域进化而成的[7]。
3.3 埋深对崖州硬皮豆出苗的影响
土壤埋藏使种子周围的环境发生一系列变化,如光照、温度、湿度、通透性以及土壤微生物等,对其萌发和出苗的调控起到关键作用。本研究结果显示,浅层的土壤埋藏为崖州硬皮豆种子萌发创造了适宜的环境条件,使出苗率得到提高。超过适宜的埋藏深度后,崖州硬皮豆种子随着埋藏深度的增加,出苗率逐渐受到抑制,其原因可能是顶土层的压力大于种子突破土层的能力[30]。除此之外,随着土壤深度的增加,土壤含氧量降低,种子在低氧分压的环境下会进入强迫休眠或吸涨后死亡[31],这也可能是导致崖州硬皮豆出苗率下降的另一个原因。Hu 等[16]认为,土壤埋藏对光强和光质的改变可能是改变长芒草出苗率的原因之一。本研究发现,崖州硬皮豆在适宜温度条件下的种子萌发对光照不敏感(图1),与土壤表层(0 cm)种子出苗率较高的结果基本吻合。埋藏深度1 cm 的种子出苗率显著高于土壤表层(P<0.05),其原因可能是表层水分蒸发过快,无法满足崖州硬皮豆这类较大粒种子萌发的水分需求,这与前人在黄花矶松(Limonium aureum)[5]和霸王(Zygophyllum xanthoxylum)[32]等植物的研究结果一致。
4 结论
本研究得出以下主要结论:1)崖州硬皮豆种子具有约32%的硬实率,在生产实践中应注意种子硬实的破除;2)崖州硬皮豆最适萌发温度在32~33 ℃,光照对崖州硬皮豆种子萌发的影响取决于温度的变化;3)随着水分和盐胁迫的加重,崖州硬皮豆种子萌发率和萌发速率呈降低的趋势,当PEG 渗透势和NaCl 浓度分别达到- 1.2 MPa和400 mmol·L-1时,种子基本不会萌发;4)相比较最终萌发率,水分和盐胁迫对崖州硬皮豆种子萌发速率的影响更为明显;5)崖州硬皮豆种子适宜播种深度为1~2 cm。
