萱草属植物种间与种内分离群体花器官主要性状的遗传分析
2019-12-19侯非凡崔虎亮王金耀邢国明亢秀萍
熊 雄,侯非凡,崔虎亮,王金耀,邢国明,李 森,亢秀萍
(1.山西农业大学园艺学院,山西太谷030801;2.山西省设施蔬菜产业提质增效协同创新中心,山西太谷030801)
黄花菜(Hemerocallis citrina Baroni)又名金针菜、柠檬萱草,属于百合科萱草属,是一种多年生草本植物[1-3]。萱草(Hemerocallis fulva)又名金针、忘忧草等,属多年生宿根草本,颜色以橘黄色为主[4]。萱草属植物在全世界有14 种,主要分布于亚洲温带和亚热带地区,我国约有11 种,是萱草属植物的自然分布中心,各个品种被广泛栽种用于园林绿化,可作观赏和食用。人工选育及创制新种质是改善现有种质资源的重要手段[5-6]。
1930 年以后,美国植物学家、园艺爱好者收集中国、日本等国家所产萱草属植物,对这些品种进行杂交育种,使得萱草属植物品种越来越丰富,现有品种已达万种以上,成为重要的观赏及切花花卉。国内外对萱草属物种细胞学、遗传学、代谢组学开展了较多的研究,主要集中在萱草属植物的栽培[3]、代谢产物的提取与优化[7-9]、染色体鉴定[5,10]、遗传多样性研究[11-12]等方面。
杂交育种是目前萱草属植物育种的主要手段,从最初的常规杂交到多种花粉混合授粉杂交,逐渐获得具有极高观赏价值的萱草品种[13-14]。杂交亲和性是杂交育种的重要内容,同时野生萱草的种质资源利用及生殖隔离的克服也变得尤为重要。PETIT研究表明,二倍体和四倍体杂交不会成功,结出的果实会在几天甚至7 d 左右后败育。国内的学者也对萱草属植物杂交亲和性进行了大量的研究。研究表明,二倍体萱草杂交亲和性因亲本的不同而存在差异,不同倍性萱草杂交的结实率低,败育现象严重[15]。萱草自然授粉自交不结实,但是部分品种人工自交结实。不同开花习性的萱草杂交试验发现,某些萱草品种之间存在生殖隔离[16]。萱草属植物中黄花菜大同与萱草摇篮曲之间存在巨大差异,亲缘关系相对较远,而黄花菜东庄与冲里在产量上具有相对的差异,所以,利用这几种材料构建种间及种内的F1分离群体,可进一步扩展萱草属植物育种资源基础[17]。
为了解析萱草属植物花器官主要性状的遗传规律,本研究利用萱草属种间杂交(大同黄花×摇篮曲)和种内杂交(东庄×冲里),对F1群体的花朵直径、花葶高度、花蕾长度、花蕾宽度、单个花蕾鲜质量等农艺性状进行测定,计算其遗传力[18],并对各农艺性状进行相关性分析,旨在得出萱草属植物的遗传方式,推动萱草属植物种质创新的进程,并为分子遗传学研究、连锁遗传图谱构建、性状QTL定位分析方面的研究提供科学依据[19-20]。
1 材料和方法
1.1 亲本材料选择
种间杂交群体选用山西省大同市黄花菜地方品种大同(P1)作母本,父本则是引种自河北保定旭东新优品种宿根花卉繁育基地的萱草摇篮曲(P2)(图1)。种内杂交群体选用山西省太谷县东庄村的黄花菜农家种东庄(P3)作母本,父本选用湖南黄花菜地方种冲里(P4)(图1)。

1.2 试验方法
1.2.1 大同黄花与摇篮曲杂交 参考侯非凡[21]的授粉方法进行父母本杂交,父本摇篮曲单朵花开花时间为每天6:00—9:00,每日早晨父本花朵开放后立即采集花粉并干燥,套袋隔离即将开放用作父本的花朵。在母本大同花朵还未开放前,每日16:00开始杂交,次日6:00—8:00 进行复授粉,花期结束后收集成熟的果子。之后进行催芽,待种子露白后,将露白的种子播种于穴盘或苗床。
1.2.2 东庄与冲里杂交 参考侯非凡[21]的授粉方法进行父母本杂交,每日6:00 采集父本植株的花粉,将花粉干燥,17:00 对第2 天将采集的父本花朵套袋。选择母本植株即将开放的花蕾,去掉花药,将父本花粉涂在去雄花蕾柱头上,套袋。次日6:00—8:00 复授粉提高杂交成功率。花期杂交结束后,10 月中下旬收集成熟果子。之后将杂交种子进行催芽,7 d 左右种子开始露白,第2 周是集中萌发期,种子萌发过程持续28 d 左右。露白的种子播种于穴盘或苗床,采用草炭∶珍珠岩∶蛭石=3∶1∶1(体积比)基质进行育苗。
1.3 测定项目及方法

表1 F1 群体性状描述
本研究对大同×摇篮曲群体、冲里×东庄群体的亲本与F1进行花朵直径、花蕾长度等方面的遗传分析,对主要的遗传性状进行相关性分析,并对亲本与F1进行形态学的测量与分析。采用游标卡尺测量花朵直径、花蕾长度、花蕾宽度等指标,采用卷尺测量花葶高度,采用千分之一天平测量单个花蕾鲜质量、干质量,具体测定标准参照表1。根据具体的目的与综合的观赏性状比较亲本与F1之间的异同,以期得到新的种质[22]。
1.4 数据分析
采用Excel 2010 软件对各数量性状进行变异系数分析。变异系数表示各个性状之间的变异程度,是标准差与平均数的比值,变异系数大表示变异的程度大[22]。
遗传力指亲本遗传给后代某一性状的能力,是遗传方差在总方差中所占的比值,可以作为杂种后代进行选择的一个指标[22]。2 个群体的遗传力计算采用SPSS 软件进行统计。

式中,H2为遗传力;Vg 为遗传变量;Vp 为表型变量。
采用R 语言来计算各个性状之间相关性,并证明2 个群体花葶高度遗传属于数量遗传。
2 结果与分析
2.1 杂交后代主要农艺性状遗传分析

表2 2 个杂交群体农艺性状遗传变异参数
大同×摇篮曲杂交得到的F1群体与东庄×冲里杂交得到的F1群体在杂交中未出现败育现象,其结实率高,出苗较为整齐,得到了足够的成苗用于后续的数据分析。通过对F1进行遗传分析得出,2 个杂交群体花朵直径、花葶高度、花蕾长度、花蕾宽度、单个花蕾鲜质量的遗传力在32.8%~86.9%。由表2 可知,摇篮曲×大同F1群体中变异幅度最小的是花蕾长度,为8.28%,变异幅度最大的是单个花蕾鲜质量,为17.43%。东庄×冲里F1群体中单个花蕾鲜质量变异系数较大,为25.43%。东庄×冲里F1群体花朵直径遗传力较高,为86.9%,而大同×摇篮曲F1群体的花朵直径遗传力较低,为32.8%,2 种杂交组合主要农艺性状的遗传力均表现为花葶高度遗传力较高,分别为73.2%,65.4%,花蕾宽度遗传力较低,分别为41.2%,34.2%。
2.1.1 花朵直径遗传变异分析 大同×摇篮曲杂交组合中花朵直径的标准差F1>摇篮曲>大同。母本为山西大同黄花,花型优美,花朵繁多,一般为蜘蛛型花朵,内外花瓣狭长,花色为黄色,开花时间为19:00 左右。因此,将其与株型低矮,花葶高度为56.85~79.36 cm、紧凑且花朵较大,花色为淡粉色,开花时间为8:00 左右的摇篮曲萱草进行杂交,以期获得花色丰富、抗性较强的新种质。从图2 可以看出,通过杂交得到了花朵直径超越父母本的植株,经过计算,F1的花朵直径遗传力为32.8%,提高了F1花朵直径的遗传变异系数,扩大了遗传变异的范围,得到了更多的变异和杂种优势。

东庄×冲里杂交组合的花朵直径标准差为F1>冲里>东庄(表2)。父本为祁东黄花菜冲里,花色为黄色,花型优美,植株高大,花葶高度为149.60~166.20 cm,单株产量较高,具有一定的倒伏性。母本为农家种东庄,植株不易倒伏,花色为黄色,花瓣呈狭长型,单株产量较高。由图3 可知,这2 种黄花菜进行杂交得到了花朵直径超越父母本的植株,为创造高产新种质提供了一定的基础。

2.1.2 花葶高度遗传变异分析 由图4 可知,大同×摇篮曲杂交组合中,母本大同黄花的花葶高度平均值最高,其次为F1,最小的是父本摇蓝曲,且标准差为母本>F1>父本。父本花葶高度较低,为69.77 cm,较稳定。母本花葶高度较高,为104.53 cm,变异系数较大,为13.93%。通过计算得出F1的花葶高度遗传力为65.4%。

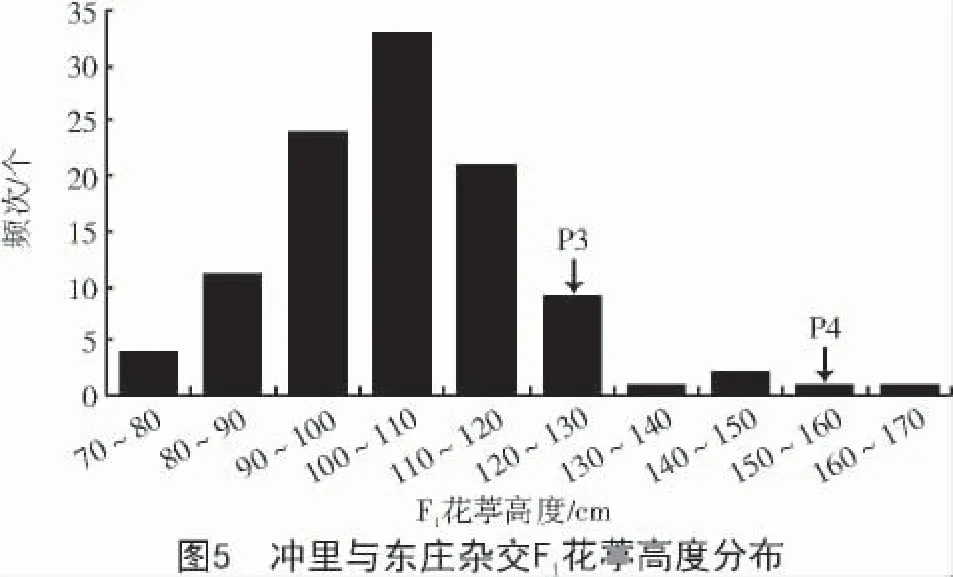
由图5 可知,对于东庄×冲里杂交组合,父本冲里花葶高度的平均值最高,F1花葶高度平均值最低,且花葶高度的标准差为F1>母本>父本,父本花葶高度在149.60~166.20 cm,母本花葶高度在110.20~139.60 cm,以期获得适合人们采摘花蕾的最佳高度。F1变异系数较大,为16.58%,最终花葶高度的遗传力为73.2%。
观察2 个F1花葶高度可看出,结果有一定的正态性,从图6 可以看出,采用R 语言中的Q-Q plot 得出花葶高度符合正态分布,遗传方式属于数量遗传。

2.1.3 花蕾长度遗传变异分析 大同×摇篮曲群体的花蕾长度发生了一定的变异,但变异程度不大,其母本大同花蕾长度平均值最高,父本最低,F1平均值则在父母本之间,标准差则为F1>母本>父本。母本花蕾长度较长且较为稳定,F1变异系数较大,为8.28%,花蕾长度遗传力为60.5%。
东庄×冲里群体中,父母本花蕾长度均在110 mm 左右,F1花蕾长度平均值最高,而标准差为F1>母本>父本,且子代与父母本相比,其花蕾长度存在超亲现象。花型均为蜘蛛型,内外花被较为狭长,F1变异系数较大,为12.56%,花蕾长度遗传力为46.2%。
2.1.4 花蕾宽度遗传变异分析 大同×摇篮曲群体的母本大同花蕾呈长条形,花蕾宽度较窄,而父本摇篮曲花蕾宽度较宽,然而其长度较短。花蕾宽度平均值为父本>F1>母本,标准差为F1>父本>母本,花蕾宽度遗传力为34.2%。
东庄×冲里群体父母本花蕾均为狭长型,花蕾宽度较为接近,且都小于萱草的花蕾宽度。其花蕾宽度平均值为F1>母本>父本,标准差为F1>母本>父本。F1的变异系数较大,为17.32%,花蕾宽度遗传力为41.2%。
2.1.5 单个花蕾鲜质量遗传变异分析 大同×摇篮曲群体,父本摇篮曲单个花蕾鲜质量较大,母本大同单个花蕾鲜质量较小,单个花蕾鲜质量平均值为父本>F1>母本,标准差为父本>F1>母本。F1的变异系数较大,为17.43%,单个花蕾鲜质量的遗传力为66.4%。
东庄×冲里群体,父本冲里与母本东庄的单个花蕾鲜质量差异不大,单个花蕾鲜质量平均值为F1>母本>父本,标准差为F1>母本>父本。F1变异系数较大,为25.43%,单个花蕾鲜质量遗传力为58.6%。
2.2 农艺性状之间的相关性分析
根据2.1 的试验结果发现,2 个群体的花朵直径、花葶高度、花蕾长度、花蕾宽度、单个花蕾鲜质量之间具有一定的相关性,对5 个农艺性状的指标进行相关性分析得出,相关性较强的性状为花蕾长度与花蕾宽度、花蕾长度与单个花蕾鲜质量、花蕾宽度与单个花蕾鲜质量[23]。

表3 大同×摇篮曲群体5 个农艺性状相关性分析
由表3 可知,大同×摇篮曲群体花蕾长度与花蕾宽度之间的相关系数为0.45,花蕾长度与单个花蕾鲜质量之间的相关系数为0.72,花蕾宽度与单个花蕾鲜质量之间的相关系数为0.65,根据Pearson 相关系数计算得出,花蕾长度与花蕾宽度呈极显著正相关,花蕾长度与单个花蕾鲜质量呈极显著正相关,花蕾宽度与单个花蕾鲜质量呈极显著正相关。其他指标之间相关性较差,呈现弱相关或不相关[22]。

表4 东庄×冲里群体5 个农艺性状相关性分析
由表4 可知,东庄×冲里群体花蕾长度与花蕾宽度之间的相关系数为0.96,花蕾长度与单个花蕾鲜质量之间的相关系数为0.98,花蕾宽度与单个花蕾鲜质量之间的相关系数为0.99,根据Pearson 相关系数计算得出,花蕾长度与花蕾宽度呈极显著正相关,花蕾长度与单个花蕾鲜质量呈极显著正相关,花蕾宽度与单个花蕾鲜质量呈极显著正相关,其他性状之间不相关[17-18]。
3 结论与讨论
杂交育种是萱草属植物丰富种质资源的主要育种手段,种间杂交育种可以将优良性状集中到杂种上,对于创造新种质、丰富变异类型、探索生物进化过程有重要的意义。花期性状作为种间杂交群体的主要观赏性状,是对观赏植物进行品质选择的必要基础。其主要目的是选择花大色艳,花期较长的种质[24]。
高超利[25]研究发现,萱草品种金娃娃与红宝的杂交后代花期变异系数小,变异系数最大的是花序着花数。对2 个大花萱草品种金娃娃与红宝杂交后代物候进行观测发现,花期的变异较小,仅有1 株花期超15 d,其他花期超亲现象不明显。朱云华[11]证实了萱草属植物种间杂交的可能性,从5 个杂交组合中选出花型、花色优异的种质资源。侯祥云等[13]对芍药组内组间杂交群体进行分析说明了组内、组间杂交的可能性。朱蕊蕊等[22,26]证实了耧斗菜种内杂交的可能性,并通过对现有种质筛选出适宜北方地区栽种的耧斗菜,进一步丰富了种质。祝朋芳等[16]对萱草属植物花柱进行不同处理,证明大花萱草品种间存在不同程度的杂交障碍。本研究表明,摇篮曲×大同种间群体的花葶高度与单个花蕾鲜质量的遗传力较高,且单个花蕾鲜质量的变异系数较大;冲里×东庄群体的单个花蕾鲜质量变异系数较大,花葶高度与花朵直径的遗传力较高;同时2 个杂交群体的花蕾长度、花蕾宽度、单个花蕾鲜质量相关性呈极显著相关;对于种间杂交群体,花蕾越长,单个花蕾鲜质量越高;对于种内杂交群体,花蕾越宽,单个花蕾鲜质量越高。赵珺[24]对萱草属植物种间杂交的花性状研究发现,影响萱草花期的因素还有很多,花期的相关性状均是由多基因控制的数量性状遗传,如花葶高度、花序长度、花蕾的长宽等。今后应对萱草的花期、花色、开花时间等性状进行遗传分析及相关性分析。表型观测进行遗传分析,明确萱草属植物的遗传模型,估计主基因的遗传效应,为萱草属植物进行选育,扩大萱草属遗传理论基础依据具有更新一步的启发[24]。
本研究对这2 个群体的观赏性状进行遗传分析,但是表型属于遗传和环境相互作用的结果,受外界环境因素影响较大,比如气候、土壤等。所以,得出的结论要进行进一步探索与研究,而细胞水平、分子水平的研究正好填补了这部分空白[24]。杂交育种的研究为选育新品种打下良好的基础,得到稳定的栽培种,为进行连锁图谱的构建及QTL 定位打下坚实的基础。
