广义李属植物叶脉序特征及其分类学意义
2019-12-18黄文鑫吴保欢石文婷羊海军崔大方
黄文鑫, 吴保欢, 石文婷, 羊海军, 崔大方,①
(1. 深圳市罗湖区市政园林管理所, 广东 深圳 518001; 2. 华南农业大学: a. 林学与风景园林学院, b. 公共基础课实验教学中心, 广东 广州 510642)
广义李属〔PrunusLinn. (s.l.)〕隶属于蔷薇科(Rosaceae)李亚科(Subfam. Prunoideae),为核果类落叶或常绿乔木、灌木,全球有200~250种,广泛分布于北半球,北美洲、欧洲和亚洲北部为主要分布地[1]401-432。广义李属植物种类众多,从茂密的森林到荒芜的沙漠,从地势平坦的平原到寒冷的高山地带均有分布[2],其中许多种类以经济价值和观赏价值高为人们熟知,例如:著名的水果桃〔Prunuspersica(Linn.) Stokes〕、李(PrunussalicinaLindl.)、梅〔Prunusmume(Sieb.) Sieb. et Zucc.〕和杏(PrunusarmeniacaLinn.),常见的干果扁桃〔Prunusdulcis(Mill.) D. A. Webb〕和山杏(PrunussibiricaLinn.),以及闻名遐迩的观赏植物东京樱花(PrunusyedoensisMatsum.)和稠李(PrunuspadusLinn.)等。
广义李属的分类可以追溯到1700年Tournefort等[3]记录的狭义李属〔PrunusLinn. (s.s.)〕、杏属(ArmeniacaScop.)、樱属(CerasusMill.)、扁桃属(AmygdalusLinn.)、桃属(PersicaMill.)和桂樱属(LaurocerasusDuhamel)6个属。1753年,Linnaeus[4]在《Species Plantarum》一书中,仅接受了AmygdalusLinn.和PrunusLinn. 2个属。此后广义李属的分类处理一直存在争议,概括而言主要集中在以下3个方面:1)李属的范围不确定,一些学者使用小属的概念,李属仅包含果实表面被蜡质、果核扁平而平滑的种类,如《中国植物志》[5]1-3和《Flora of China》[1]401-432,部分学者则采用更宽泛的李属概念,如Bentham等[6]就将此前的杏属、樱属、扁桃属和桂樱属等归到广义李属中,Kalkman[7]和Wen等[8]也分别将臀果木属(PygeumGaertn.)和臭樱属(MaddeniaHook. f. et Thoms.)归到广义李属中;2)属下类群分类处理迥异,如Rehder[9]在樱亚属(Subg.CerasusA. Gray)下设矮樱组(Sect.MicrocerasusC. K. Schneid.),而Focke[10]则将矮樱组和樱属处理为李属的亚属等级;3)部分种的分类处理仍然存在争议,如斑叶稠李(PrunusmaackiiRupr.)有时被划分至樱亚属[11],有时则被划分至稠李亚属〔Subg.Padus(Moench) Koehne〕[5]94,而Chen等[12]描述的新种磐安樱(PrunuspananensisZ. L. Chen, W. J. Chen et X. F. Jin)则被王贤荣[13]3认为和尾叶樱桃(PrunusdielsianaC. K. Schneid.)是同一分类实体。
叶脉是植物叶片中的维管束组织成分,在叶片中彼此连通,形成一个有规律的网状结构[14]。叶脉序能稳定遗传,具有特定的排列模式,且这一模式在幼叶阶段已经形成,不易受环境影响,是研究植物分类学与植物系统学的重要工具[15]。叶脉在科间、属间乃至种间都有重要的分类学意义,对于属间和种的划分也有重要的参考价值[16-17],但目前国内外尚未见有关广义李属植物叶脉序的研究。为此,本文研究了广义李属70种(含67种和3变种,下同)植物的叶脉序特征,通过光学显微镜对植物叶脉序特征进行观察和比较,寻求有分类意义的叶脉序性状,旨在为进一步研究广义李属植物的分类和系统演化提供数据。
1 材料和方法
于2017年1月至6月从华南农业大学植物标本馆(CANT)馆藏广义李属70种植物的腊叶标本(表1)上取成熟叶片,每份标本取3枚。参考Ellis等[18]210-211的方法制作叶脉观察玻片,制作流程为:根据种类的不同,将成熟叶片放入质量体积分数5%NaOH溶液中静置1~36 h;待叶肉细胞离析后取出,用毛刷扫去叶表皮和叶肉组织;流水清洗后用体积分数50%、75%和95%乙醇梯度脱水,制成临时装片,在Olympus C23光学显微镜(日本Olympus公司)下观察叶片最宽处区域并拍照。
表1 广义李属植物凭证标本信息
Table 1 Voucher specimens ofPrunusLinn. (s.l.)

编号No.种(变种)Species (Variety)采集人Collector凭证标本号No. of voucher specimen采集地Locality采集日期Collection date(CCYY-MM-DD)李亚属Subg. Prunus1杏Prunus armeniaca崔大方等CUI Dafang, et alPP2016026中国云南丽江Lijiang of Yunnan in China2016-05-312藏杏Prunus armeniaca var. holosericea崔大方等CUI Dafang, et alPP2016204中国西藏昌都Qamdo of Tibet in China2016-05-313梅Prunus mume崔大方等CUI Dafang, et alPP2016027中国云南丽江Lijiang of Yunnan in China2016-05-314山杏Prunus sibirica无采集人Anonymus20150709005中国内蒙古呼伦贝尔Hulun Buir of Inner Mongolia in China2015-07-095东北杏Prunus mandshurica崔大方等CUI Dafang, et alPP2015553中国黑龙江哈尔滨Harbin of Heilongjiang in China2015-06-056政和杏Prunus zhengheensis吴保欢WU BaohuanFJZH2014009中国福建南平Nanping of Fujian in China2014-11-187李梅杏Prunus limeixing吴保欢WU BaohuanLNXY2014001中国辽宁营口Yingkou of Liaoning in China2014-10-288李Prunus salicina崔大方等CUI Dafang, et alPP2015464中国黑龙江哈尔滨Harbin of Heilongjiang in China2015-06-029欧洲李Prunus domestica崔大方等CUI Dafang, et alPP2015868中国新疆伊犁Ili of Xinjiang in China2015-07-0210樱桃李Prunus cerasifera崔大方等CUI Dafang, et alPP2015780中国新疆伊犁Ili of Xinjiang in China2015-06-3011黑刺李Prunus spinosa许可望XU KewangXKW-254俄罗斯莫斯科Moscow in Russia2016-09-0812海滩李Prunus maritima吴保欢WU BaohuanAP-003美国马萨诸塞波士顿Boston of Massachusetts in USA2015-10-02桃亚属Subg. Amygdalus13桃Prunus persica吴保欢WU BaohuanPP-10067中国陕西西安Xian of Shaanxi in China2014-06-0114山桃Prunus davidiana吴保欢WU BaohuanPP2015608中国新疆乌鲁木齐Urumqi of Xinjiang in China2015-06-2515甘肃桃Prunus kansuensis吴保欢WU BaohuanPP-10069中国陕西宝鸡Baoji of Shaanxi in China2014-06-0216光核桃Prunus mira崔大方等CUI Dafang, et alPP2016097中国云南香格里拉Shangri-La of Yunnan in China2016-06-0117蒙古扁桃Prunus mongolica朝落门CHAO Luomen462中国内蒙古阿拉善Alagxa of Inner Mongolia in China2016-05-1718西康扁桃Prunus tangutica吴保欢WU BaohuanPP-10011中国甘肃甘南Gannan of Gansu in China2014-05-2519榆叶梅Prunus triloba崔大方等CUI Dafang, et alPP2015668中国新疆塔城Tacheng of Xinjiang in China2015-06-29稠李亚属Subg. Padus20Prunus serotina许可望XU KewangXKW-252俄罗斯莫斯科Moscow in Russia2016-09-0821斑叶稠李Prunus maackii 许可望XU KewangXKW-250俄罗斯莫斯科Moscow in Russia2016-09-0822稠李Prunus padus许可望XU KewangXKW-248俄罗斯莫斯科Moscow in Russia2016-09-0823粗梗稠李Prunus napaulensis吴保欢等WU Baohuan, et alPP2015417中国云南丽江Lijiang of Yunnan in China2015-05-2524短梗稠李Prunus brachypoda吴保欢WU BaohuanPP-10065中国陕西西安Xi’an of Shannxi in China2015-06-0125灰叶稠李Prunus grayana吴保欢WU BaohuanLXP-X3-05689中国江西吉安Ji’an of Jiangxi in China2014-04-1626细齿稠李Prunus obtusata吴保欢WU BaohuanPHS1001中国四川阿坝Aba of Sichuan in China2014-06-1827橉木Prunus buergeriana吴保欢WU BaohuanDWS2014043中国湖南长沙Changsha of Hu’nan in China2014-07-29桂樱亚属Subg. Laurocerasus28Prunus caroliniana吴保欢WU BaohuanAP-004美国加利福尼亚戴维斯Davis of California in USA2015-10-1529Prunus ilicifolia吴保欢WU BaohuanAP-008美国加利福尼亚戴维斯Davis of California in USA2015-10-1530葡萄牙桂樱Prunus lusitanica吴保欢WU BaohuanAP-009美国加利福尼亚戴维斯Davis of California in USA2015-10-1531桂樱Prunus laurocerasus许可望XU KewangXKW-258俄罗斯莫斯科Moscow in Russia2016-09-0832刺叶桂樱Prunus spinulosa陈雪梅CHEN XuemeiPL1029中国广东梅州Meizhou of Guangdong in China2014-10-0133大叶桂樱Prunus zippeliana陈雪梅CHEN XuemeiPL1028中国广东梅州Meizhou of Guangdong in China2014-10-0134尖叶桂樱Prunus undulata陈雪梅CHEN XuemeiPL1009中国广东韶关Shaoguan of Guangdong in China2014-08-1835坚核桂樱Prunus jenkinsii陈雪梅CHEN XuemeiPL1013中国云南西双版纳Sipsongpanna of Yunnan in China2014-09-0436长叶桂樱Prunus dolichophylla陈雪梅等CHEN Xuemei, et alPL1019中国云南西双版纳Sipsongpanna of Yunnan in China2014-09-0437腺叶桂樱Prunus phaeosticta陈雪梅CHEN XuemeiPL1010中国广东惠州Huizhou of Guangdong in China2014-08-2138南方桂樱Prunus austrosinensis王盼WANG PanPL1027中国广东清远Qingyuan of Guangdong in China2014-09-12
叶脉序特征描述参考Ellis等[18]1-70的叶脉结构特征术语体系,选取其中8个性状(图1)用于广义李属植物的叶脉序特征描述,并对各性状进行赋值。1)二级脉(侧脉):达缘脉(二级脉延伸至叶缘),赋值1;半达缘脉(二级脉于近叶缘处分支,一条达缘,另一条连于上端相邻二级脉),赋值2;花环状半达缘脉(二级脉有多组环状结构,最外侧环的分支脉达缘),赋值3;简单弓形脉(二级脉相连形成显著的弓形或环状结构),赋值4。2)细二级脉:有,赋值1;无,赋值2。3)间二级脉:无,赋值1;每个二级脉区间少于1条,赋值2;每个二级脉区间通常为1条,赋值3;每个二级脉区间多于1条,赋值4。4)二级脉间三级脉:对生贯穿(大多数三级脉平行穿越于相邻的2条二级脉间,且不分支),赋值1;混合贯穿(三级脉同时存在对生和互生的贯穿脉,互生贯穿脉是指大多数穿越二级脉的三级脉在二级脉间区中央处有较规则的分支),赋值2;网状(三级脉与其他三级脉或二级脉连结,组成网格状结构),赋值3。5)四级脉:规则网状(四级脉与其他叶脉相互连结成网格状结构,且叶脉相交的角度规则),赋值1;不规则网状(四级脉与其他叶脉相互连接成网格状结构,且叶脉相交的角度变化较大),赋值2;6)五级脉:规则网状(五级脉与其他叶脉相互连结成网格状结构,且叶脉相交的角度规则),赋值1;不规则网状(五级脉与其他叶脉相互连接成网格状结构,且叶脉相交的角度变化较大),赋值2;自由分支(五级脉自由分支,为叶片上最细的脉序),赋值3。7)脉间区(叶脉围成的最小叶组织单元)发育:差(脉间区具有7条以上的边,形状大小不规则),赋值1;中(脉间区形状不规则,大小略有变化),赋值2;良(脉间区形状大小较一致,一般具有3~6条边),赋值3。8)游离端小脉(自由分支的最高次脉):不分支,赋值1;简单分支,赋值2;二歧分支,赋值3;不均等分支,赋值4。
2 结果和分析
供试广义李属70种植物的叶脉序特征存在丰富的变异(图版Ⅰ、图版Ⅱ和表2),但具有一定的共性,表现为一级脉仅中脉1条;二级脉多为半达缘脉,少数为花环状半达缘脉或简单弓形脉;二级脉间三级脉和四级脉不分支;脉间区发育,由三级脉和四级脉构成;具游离端小脉。
表2 广义李属植物叶脉序特征
Table 2 Leaf venation characteristics ofPrunusLinn. (s.l.)

编号 No.种(变种) Species (Variety)SV1)MSV2)IV3)ITV4)QuaV5)QuiV6)AD7)FEV8)李亚属Subg. Prunus1杏Prunus armeniaca312212222藏杏Prunus armeniaca var. holosericea312212313梅Prunus mume312212224山杏Prunus sibirica312211315东北杏Prunus mandshurica312211316政和杏Prunus zhengheensis322111317李梅杏Prunus limeixing313212218李Prunus salicina323211319欧洲李Prunus domestica3222113110樱桃李Prunus cerasifera3222113111黑刺李Prunus spinosa3222113112海滩李Prunus maritima32221221桃亚属Subg. Amygdalus13桃Prunus persica2232222214山桃Prunus davidiana2232222315甘肃桃Prunus kansuensis2242221416光核桃Prunus mira2232222317蒙古扁桃Prunus mongolica2222222318西康扁桃Prunus tangutica2222222219榆叶梅Prunus triloba22222221稠李亚属Subg. Padus20Prunus serotina2242222321斑叶稠李Prunus maackii 2222222322稠李Prunus padus3222222323粗梗稠李Prunus napaulensis3232222324短梗稠李Prunus brachypoda3222222225灰叶稠李Prunus grayana3222222226细齿稠李Prunus obtusata3222222227橉木Prunus buergeriana22422223桂樱亚属Subg. Laurocerasus28Prunus caroliniana4232222329Prunus ilicifolia3222222330葡萄牙桂樱Prunus lusitanica2213222431桂樱Prunus laurocerasus3222222332刺叶桂樱Prunus spinulosa3232222333大叶桂樱Prunus zippeliana2212222334尖叶桂樱Prunus undulata4233222435坚核桂樱Prunus jenkinsii2242222336长叶桂樱Prunus dolichophylla2232222337腺叶桂樱Prunus phaeosticta4233222338南方桂樱Prunus austrosinensis22322314樱亚属Subg. Cerasus39川西樱桃Prunus trichostoma2222222140雕核樱桃Prunus pleiocerasus2222222241欧洲甜樱桃Prunus avium 2222113242华中樱桃Prunus conradinae 2221113143多毛樱桃Prunus polytricha2212123244微毛樱桃Prunus clarofolia2212123245高盆樱桃Prunus cerasoides2221113146麦李Prunus glandulosa2222222247毛樱桃Prunus tomentosa1222222148尾叶樱桃Prunus dielsiana2221122249磐安樱Prunus pananensis22211222
1)SV: 二级脉Secondary vein. 1: 达缘脉Craspedodromous; 2: 半达缘脉Semicraspedodromous; 3: 花环状半达缘脉Festooned semicraspedodromous; 4: 简单弓形脉Simple brochidocromous.
2)MSV: 细二级脉Minor secondary vein. 1: 有Present; 2: 无Absent.
3)IV: 间二级脉Intersecondary vein. 1: 无Absent; 2: 每个二级脉区间少于1条Intersecondary vein per intercostal area is less than one; 3: 每个二级脉区间通常为1条Intersecondary vein per intercostal area is usually one; 4: 每个二级脉区间多于1条Intersecondary veins per intercostal area are more than one.
4)ITV: 二级脉间三级脉Intercostal tertiary vein. 1: 对生贯穿Opposite percurrent; 2: 混合贯穿Mixed percurrent; 3: 网状Reticulate.
5)QuaV: 四级脉Quaternary vein. 1: 规则网状Regular reticulate; 2: 不规则网状Irregular reticulate.
6)QuiV: 五级脉Quinternary vein. 1: 规则网状Regular reticulate; 2: 不规则网状 Irregular reticulate; 3: 自由分支Freely ramifying.
7)AD: 脉间区发育Areolation development. 1: 差Poor; 2: 中Modetate; 3: 良Good.
8)FEV: 游离端小脉Freely ending veinlet. 1: 不分支Unbranched; 2: 简单分支Simple branched; 3: 二歧分支Dichotomous branching; 4: 不均等分支Dendritic branching.
2.1 二级脉、细二级脉和间二级脉类型
广义李属植物的二级脉分为达缘脉、半达缘脉、花环状半达缘脉和简单弓形脉(表2和图2),以半达缘脉居多,其中,李亚属(Subg.PrunusLinn.)和稠李亚属多为花环状半达缘脉,桃亚属(Subg.AmygdalusLinn.)和樱亚属多为半达缘脉,桂樱亚属(Subg.LaurocerasusKoehne)为半达缘脉、花环状半达缘脉和简单弓形脉,臀果木类群(Pygeumgroup)为简单弓形脉,臭樱类群(Maddeniagroup)为达缘脉和半达缘脉。
广义李属植物中,仅李亚属中的杏、藏杏(Prunusarmeniacavar.holosericeaBatalin)、梅、山杏、东北杏〔Prunusmandshurica(Maxim.) Koehne〕和李梅杏(PrunuslimeixingJ. Y. Zhang et M. N. Lu)有细二级脉,其他广义李属植物均无细二级脉。
广义李属植物中,李亚属、桃亚属和桂樱亚属植物每个二级脉区间的间二级脉多为1条或少于1条,樱亚属植物少于1条或无,臀果木类群和臭樱类群植物无间二级脉。

1. 四川臭樱,二级脉为达缘脉Prunus hypoxantha (Koehne) J. Wen, secondary vein is craspedodromous; 2. 坚核桂樱,二级脉为半达缘脉Prunus jenkinsii Hook. f., secondary vein is semicraspedodromous; 3. 李,二级脉为花环状半达缘脉Prunus salicina Lindl., secondary vein is festooned semicraspedodromous; 4. 臀果木,二级脉为简单弓形脉Pyguem topengii Merr., secondary vein is simple brochidodromous.图2 广义李属植物二级脉类型Fig. 2 Type of secondary veins of Prunus Linn. (s.l.)
2.2 二级脉间三级脉、四级脉和五级脉类型
广义李属植物的二级脉间三级脉分为对生贯穿、混合贯穿和网状(表2和图3),且多为混合贯穿,其中,政和杏(PrunuszhengheensisJ. Y. Zhang et M. N. Lu)、华中樱桃(PrunusconradinaeKoehne)、高盆樱桃(PrunuscerasoidesD. Don)、尾叶樱桃、磐安樱、东京樱花、臀果木(PygeumtopengiiMerr.)和云南臀果木(PygeumhenryiDunn.)为对生贯穿,而葡萄牙桂樱(PrunuslusitanicaLinn.)、尖叶桂樱(PrunusundulataBuch.-Ham. ex D. Don)、腺叶桂樱〔Prunusphaeosticta(Hance) Maxim.〕和长尾毛柱樱桃〔Prunuspogonostylavar.obovata(Koehne) F. P. Metcalf〕为网状,其他供试广义李属植物为混合贯穿。

1. 高盆樱桃,二级脉间三级脉为对生贯穿Prunus cerasoides D. Don, intercostal tertiary vein is opposite percurrent; 2. 稠李,二级脉间三级脉为混合贯穿Prunus padus Linn., intercostal tertiary vein is mixed percurrent; 3. 尖叶桂樱,二级脉间三级脉为网状Prunus undulata Buch.-Ham. ex D. Don, intercostal tertiary vein is reticulate.图3 广义李属植物二级脉间三级脉类型Fig. 3 Type of intercostal tertiary veins of Prunus Linn. (s.l.)
广义李属植物的四级脉和五级脉主要分为规则网状和不规则网状,仅南方桂樱(PrunusaustrosinensisHuan C. Wang)的五级脉为自由分支(表2和图4)。李亚属和臭樱类群的四级脉为规则网状,桃亚属、稠李亚属、桂樱亚属和臀果木类群为不规则网状,樱亚属则为规则网状和不规则网状。桃亚属、稠李亚属、桂樱亚属(南方桂樱除外)、臀果木类群和臭樱类群的五级脉为不规则网状,李亚属和樱亚属为规则网状和不规则网状。

1. 东北杏,四级脉和五级脉为规则网状Prunus mandshurica (Maxim.) Koehne, quaternary and quinternary veins are regular reticulate; 2. 灰叶稠李,四级脉和五级脉为不规则网状Prunus grayana Batalin, quaternary and quinternary are veins irregular reticulate; 3. 南方桂樱,四级脉为不规则网状,五级脉为自由分支 Prunus austrosinensis Huan C. Wang, quaternary vein is irregular reticulate and quinternary vein is freely ramifying.图4 广义李属植物四级脉和五级脉类型Fig. 4 Types of quaternary veins and quinternary veins of Prunus Linn. (s.l.)
2.3 脉间区发育和游离端小脉类型
广义李属植物的脉间区发育分为差、中和良(表2和图5),其中,李亚属和樱亚属为中或良,桃亚属、稠李亚属、桂樱亚属、臀果木类群和臭樱类群多为中。
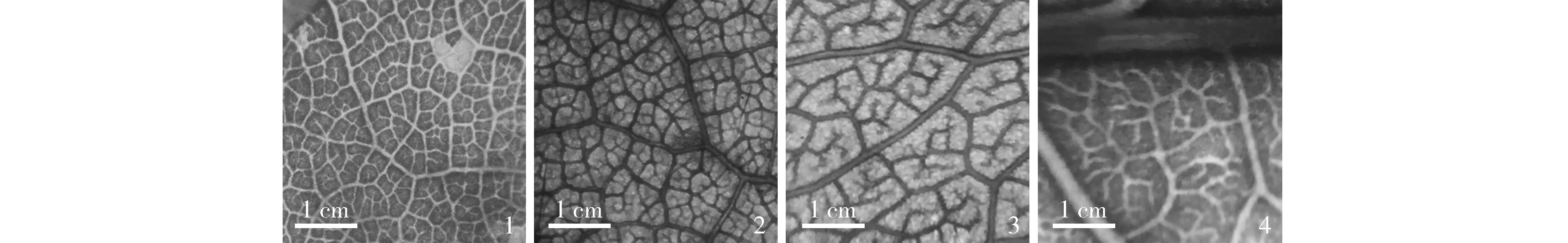
1. 东北杏,脉间区发育良,游离端小脉不分支Prunus mandshurica (Maxim.) Koehne, areolation development is good, freely ending veinlets are unbranched; 2. 迎春樱,脉间区发育中,游离端小脉简单分支Prunus discoidea (T. T. Yu et C. L. Li) Z. Wei et Y. B. Chang, areolation development is moderate, freely ending veinlets are simple branched; 3. Prunus serotina Ehrh.,游离端小脉发育中,游离端小脉二歧分支Prunus serotina Ehrh., areolation development is moderate, freely ending veinlets are dichotomous branching; 4. 南方桂樱,脉间区发育差,游离端小脉不均等分支Prunus austrosinensis (Yü et Lu) Q. H. Chen, areolation development is poor, freely ending veinlets are dendritic branching.图5 广义李属植物脉间区发育和游离端小脉类型Fig. 5 Types of areolation development and freely ending veinlets of Prunus Linn. (s.l.)
广义李属植物的游离端小脉分为不分支、简单分支、二歧分支和不均等分支(表2和图5)。李亚属多为不分支,桃亚属为不分支、简单分支、二歧分支和不均等分支,稠李亚属为简单分支和二歧分支,桂樱亚属为二歧分支和不均等分支,樱亚属植物为不分支、简单分支和二歧分支,臀果木类群为简单分支和二歧分支,臭樱类群不分支和简单分支。
3 讨 论
3.1 叶脉序研究在属内关系上的应用
矮樱组在广义李属中的分类地位存在争议,一些学者将其处理为樱亚属(或樱属)的下级分类群[5]41-43,另一些学者将其处理为与樱亚属并列的分类群[10]。杂交实验中,矮樱组植物与李亚属和部分桃亚属植物可以杂交,与典型樱类植物杂交则通常以失败告终,表明矮樱组与典型樱类关系较远[19]。Shi等[20]和王鹏飞等[21]分别利用多基因片段联合和RAPD分子标记的研究结果也显示:矮樱组与典型樱类的关系较远。本研究中,矮樱组植物与典型樱类植物的叶脉性状存在较明显差异,其中麦李(PrunusglandulosaThunb.)、长尾毛柱樱桃、天山樱桃(Prunusprostratavar.concolorLipsky)和毛樱桃(PrunustomentosaC. P. Thunb. ex A. Murray)的四级脉和五级脉等结构与桃亚属植物更为接近,但毛樱桃的二级脉与前三者不同,为达缘脉,这与王鹏飞等[21]基于RAPD分子标记的聚类分析结果相吻合。但在王鹏飞等[21]的聚类分析结果中,欧李(PrunushumilisBge.)和麦李组成的分支并未与桃亚属植物聚为一支,这与叶脉性状暗示的亲缘关系不同。综合来看,传统分类学意义上的樱亚属并非一个单系,矮樱组移出樱亚属是有必要的,同时矮樱组可能不是一个单系类群,有必要通过进一步的研究来确定其分类处理。
李属(PrunusLinn.)和杏属(ArmeniacaMill.)常被不同的学者处理为独立的属或亚属[5]1-25,也有学者将二者合并为李亚属,下设李组(Sect.PrunusBenth et Hook.)和杏组〔Sect.Armeniaca(Mill.) K. Koch〕[22]。与其他类群相比,李属和杏属在形态上更为近似,这2个属植物的幼叶多为席卷式,侧芽单生,花常单生,核常光滑或有不明显孔穴。此外,这2个属植物容易发生杂交,李梅杏被认为是李和杏的杂交种[23],紫杏(PrunusdasycarpaEhrh.)则是由梅和PrunusbrigantiacaVill.杂交而来[24]。众多分子研究结果也表明,在广义李属中,李属和杏属关系最为密切[11]。在叶脉结构上,李属和杏属较为相似,二级脉皆为花环状半达缘脉,四级脉为规则网状,五级脉为规则网状或不规则网状,脉间区发育中或良。因此,本研究支持将二者合并为李亚属的分类处理。
3.2 叶脉序研究在种间关系中的应用
贡山臭樱(Maddeniahimalaicavar.glabrifoliaH. Hara)与喜马拉雅臭樱(PrunushimalayanaJ. Wen)形态近似,但因其叶背近无毛作为后者的变种[25]。《Flora of China》[1]432-434将贡山臭樱并入喜马拉雅臭樱,并将喜马拉雅臭樱描述为叶背密被毛或无。Wen等[26]则认为,贡山臭樱基部心形至近心形,叶背近无毛可与四川臭樱和喜马拉雅臭樱区分,叶基部腺体多于15对可与臭樱区分,将其作为新种PrunusgongshanensisJ. Wen发表。根据观察标本,叶基部心形的性状并非贡山臭樱独有,在喜马拉雅臭樱中同样存在,且叶基部腺体常15对以上,该性状也与喜马拉雅臭樱相同。在二级脉结构上,喜马拉雅臭樱和贡山臭樱均为半达缘脉,而其他臭樱类植物为达缘脉,主要区别在于叶背有无被毛,而包括叶脉在内的特征并无明显差别,可见将贡山臭樱独立为新种不合适。因此,本研究不支持将喜马拉雅臭樱和贡山臭樱合并,建议将贡山臭樱作为喜马拉雅臭樱的变种处理。
磐安樱是Chen等[12]在浙江磐安采集到的樱类植物,其形态与浙闽樱(PrunusschneiderianaKoehne)近似,区别在于前者是苞片、子房和果实1毛,花瓣长8~9 mm,雄蕊28~30枚。磐安樱与迎春樱〔Prunusdiscoidea(T. T. Yu et C. L. Li) Z. Wei et Y. B. Chang〕也相似,但前者叶柄密被柔毛,叶基部圆形或心形,叶背面沿脉密被黄棕色柔毛。基于DNA条形码的研究结果也显示:磐安樱与浙闽樱、迎春樱、山樱花(PrunusserrulataLindl.)存在明显差异,因而被发表为新种[12]。根据叶脉研究结果,磐安樱和尾叶樱桃的叶片形态高度相似,包括叶柄和叶片密被毛和叶基部圆形等性状,且苞片、子房和果实1毛等特征也与尾叶樱桃近似,叶脉结构也无明显区别,但Chen等[12]的研究中缺少与尾叶樱桃的对比。王贤荣[13]3将磐安樱作为尾叶樱桃在浙江省的新分布,本研究支持该观点。
3.3 叶脉序多样性及其演化趋势
广义李属植物的叶脉形态具有丰富的变异,四级脉、五级脉、脉间区发育和游离端小脉存在不同程度的分化。
结合地理分布可以看出,分布于温带的落叶类群(李亚属、桃亚属、稠李亚属、樱亚属和臭樱类群)的脉间区发育以良和中为主,少数为差,游离端小脉主要为不分支、简单分支和二歧分支,少数为不均等分支;分布于亚热带-热带的常绿类群(桂樱亚属和臀果木类群)脉间区发育为中或差,游离端小脉以二歧分支为主,少数为简单分支。
现存广义李属植物种类最多的地区为亚洲东部,最早的广义李属植物化石也出土于此[27]。基于Bayes-DIVA方法,推测广义李属植物可能起源于5 800万年前的东亚地区,此后受古新世-始新世极热事件(Paleocene-Eocene Thermal Maximum)以及印度板块和欧亚板块碰撞等事件的共同驱动而快速分化。推测广义李属植物由北温带向南方的亚热带-热带迁移,叶脉密度逐渐减小,游离端小脉分支逐渐增加,其中,稠李亚属和桂樱亚属为广义李属植物从温带类型向热带类型演化的过渡类群,特别是桂樱亚属中存在许多演化程度不同的过渡种。温带地区的广义李属植物的叶片偏小,呈纸质,四级脉和五级脉较密,游离端小脉较少。而亚热带-热带地区的桂樱亚属和臀果木类群为叶片较大的全缘革质叶,四级脉和五级脉不规则且较稀疏,游离端小脉较多。
4 结 论
研究证明:叶脉序性状对广义李属植物分类及系统演化关系具有重要意义。在亚属水平上,各类群的叶脉结构具有一定的共性,如细二级脉、间二级脉、四级脉和五级脉类型等都有各自的显著特征。在种水平上,二级脉间三级脉、脉间区发育和游离端小脉等方面存在种间差别。同时,广义李属植物叶脉结构多样,具有一定的连续性,其中脉间区发育和游离端小脉呈现出不同程度的演变,可作为讨论内部演化的依据。
致谢:野外标本采集过程中得到华南农业大学俞新华老师、中山大学许可望博士、大盘山国家级自然保护区陈子林高级工程师和王盼工程师、福建武夷山国家级自然保护区李冬静工程师以及辽宁省果树科学研究所章秋平研究员等的帮助,在此一并感谢!
