芸薹属栽培种细胞质的分子标记鉴别
2019-12-16梁龙兵香2喻时周皮明雪张敏琴王仙萍赵继献
梁龙兵, 林 香2, 喻时周, 皮明雪, 张敏琴, 王仙萍, 向 阳, 赵继献
(1.贵州省农业科学院油菜研究所, 贵阳 550008; 2.贵州大学农学院, 贵阳 550025)
芸薹属(Brsssica species)是十字花科下非常重要的一个属,具有较大的经济价值,包括许多重要的油料、蔬菜和饲料作物。芸薹属栽培种即U氏三角中的6个种:3个二倍体基本种白菜型油菜(B.rapaL.或B.campestrisL.,AA,2 n=20)、甘蓝(B.oleracea, CC,2 n=18)和黑芥(B.nigra(L.) Koch,BB,2 n=16),3个四倍体复合种甘蓝型油菜(B.napusL.,AACC,2 n=38)、芥菜型油菜(B.junceaL.,Czern & Coss, AABB,2 n=36)和埃塞俄比亚芥(B.carinataA.Braun, BBCC,2 n=34)[1]。甘蓝型油菜起源于白菜型油菜和甘蓝的自然杂交及染色体加倍[2],是全世界栽培面积最为广泛的栽培种,在过去的几十年中,利用其它5个栽培种的一系列重要农艺性状极大地改良了甘蓝型油菜[3]。
植物细胞质DNA分布于线粒体和叶绿体中,由细胞质遗传系统决定,细胞质基因与细胞核基因在遗传表达中相互有信息交流[4-5],通过核质互作影响育性[6],细胞质效应对杂种优势的发挥也具有影响[7]。甘蓝型油菜的长期育种选择,使得遗传基础变窄,进行芸薹属种间远缘杂交能有效拓宽油菜遗传基础,也是油菜品种改良的重要手段之一。长期的远缘杂交,甘蓝型油菜具有来源于不同栽培种的细胞质类型,因此准确区分不同细胞质类型对于油菜杂种优势的高效利用有着重要意义[8]。由于不同物种细胞质遗传物质存在差异,芸薹属6个栽培种细胞质雄性不育类型各异[9]。研究表明,芸薹属不同物种之间,如甘蓝、芥菜型油菜、不同细胞质类型的甘蓝型油菜、白菜型油菜及黑芥、埃塞俄比亚芥、芸芥的叶绿体和线粒体基因存在较丰富的遗传多样性[13],芸薹属栽培种内如甘蓝的细胞质叶绿体基因也具有较高的多样性[10-12]。针对芸薹属6个栽培种细胞质的多样性,可以利用分子标记对不同来源的细胞质进行有效区分鉴定。本研究利用芸薹属栽培种的线粒体和叶绿体全基因组序列设计的分子标记,对来自不同地区39份芸薹属6个栽培种细胞质进行区分,为油菜材料创新以及杂种优势利用提供分子鉴定方法。
1 材料与方法
1.1 试验材料
供试材料为39份源自国内外的芸薹属不同栽培种细胞质类型的种质资源,其中白菜1份,黑芥1份,甘蓝2份,埃塞俄比亚芥1份,甘蓝型油菜26份,芥菜型油菜8份(表1)。
表1 供试材料信息

序号材料名称细胞质类型材料来源1GRB094白菜贵州省2GRB4107黑芥贵州省3GRA113甘蓝贵州省411HB32甘蓝型油菜四川省5GRB4106埃塞俄比亚芥四川省6Zhongshuang11甘蓝型油菜中国农科院油料所7GRB999甘蓝型油菜贵州省8Xiangyou17甘蓝型油菜湖南省9Chuanyou36甘蓝型油菜四川省10Y335甘蓝型油菜贵州省11Y336甘蓝型油菜贵州省12D3361A甘蓝型油菜贵州省13D3361甘蓝型油菜贵州省14AB450甘蓝型油菜澳大利亚15Liraglu甘蓝型油菜澳大利亚1631048甘蓝型油菜湖北恩施17Nabo甘蓝型油菜法国18HJa82470甘蓝型油菜芬兰19Koolzaand甘蓝型油菜荷兰20Huayou-2H甘蓝型油菜华中农业大学21Canada-2H甘蓝型油菜加拿大22TRIVMPH甘蓝型油菜加拿大23P11甘蓝型油菜捷克24Halleyuche甘蓝型油菜南朝鲜25Dong-Hae23甘蓝型油菜日本26Qinseng甘蓝型油菜日本27SV.Juno甘蓝型油菜瑞典28BF10(A)甘蓝型油菜台湾省29P4甘蓝型油菜西德30Expander甘蓝型油菜西德31Superlati-velot甘蓝型油菜新西兰32GRA093芥菜型油菜陕西33GRA094芥菜型油菜陕西34GRA096芥菜型油菜陕西35C30707芥菜型油菜贵州普安36C30854芥菜型油菜贵州安龙37C30947芥菜型油菜贵州平坝38C31085芥菜型油菜贵州织金39C31190芥菜型油菜贵州安顺
1.2 试验方法
1.2.1引物信息
根据文献资料[14-16],针对叶绿体和线粒体中的目标基因,利用39对特异性引物进行实验(表2)。
1.2.2DNA的提取
每份材料于苗期随机选取10株,采摘幼嫩叶片混合取样,根据改良CTAB法提取油菜叶片细胞质DNA[18]。经紫外分光光度计检测质量浓度后,将DNA样品浓度稀释至35 mg·μL-1,于-20 ℃保存备用。
1.2.3PCR反应
PCR检测所用的反应体系为10μL:其中模板2μL,正向引物和反向引物引物各0.5μL,TIANGEN公司的1× Taq buffer(已添加Mg+)1μL,dNTPs 0.2μL,Taq DNA聚合酶0.1μL, ddH2O 5.7μL。扩增程序: 94 ℃预变性3 min;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸30 s,35个循环,72 ℃延伸2 min。在PTC-225扩增仪上进行PCR扩增产物电泳时总电压为150 V,通常电泳1.5 h左右。扩增产物在8%非变性聚丙烯酰胺凝胶(100 mL聚丙烯酰胺溶液中含7.6 g丙烯酰胺和0.4 g甲叉双丙烯酰胺)上进行电泳分离,然后进行银染,银染方法参照《分子克隆实验指南,第三版》进行银染,在凝胶自动成像仪上观察。
1.2.4数据分析
统计电泳图谱上有差异且易于识别的多态性条带图谱上有带计为“1”,无带计为“0”,多态性信息含量(PIC)计算公式为:
式中,pi表示第i个引物的等位位点频率。用NTSYS-PC 2.10 e软件包计算遗传相似系数,获得相似系数矩阵,用该软件包的SHAN程序和UPGMA方法进行聚类分析。
2 结果与分析
2.1 细胞质SSR标记的多态性
对叶绿体、线粒体特异性引物在39份材料中的扩增结果表明,筛选到11对多态性引物,从8对线粒体特异性引物中筛选得到2对具有多态性的引物,这些多态性引物共检测到38条多态性带,每个位点的等位基因为2~6个,平均为3个,多态性信息量PIC值为0.10~0.76,平均为0.39(表3),多态性信息量衡量基因变异程度高低,当PIC≥0.5 时,该引物为高度多态性信息引物,当 0.25≤PIC<0.5 时为中度多态性信息引物,当PIC<0.25 时为低度多态性信息引物。11对多态性引物中有5对引物为高度多态性信息引物,其中包含2对线粒体引物,表明该5个特异性引物能较好地鉴别6个芸薹属栽培种。
叶绿体特异性引物PsbB-PsbT检测3个多态性类型(图1 A),在甘蓝型、芥菜型油菜栽培种内均有多态性,其中日本与朝鲜甘蓝型油菜在片段270 bp、320 bp同时出现2个多态类型,表明不能完全区分2种芸苔属栽培种。图1 B为线粒体引物BnTR 1可得3个多态性类型(图1 B),在甘蓝型、芥菜型油菜栽培种内均有2种多态类型。其中加拿大、捷克甘蓝型油菜出现多态性,贵州安顺与织金出现多态类型。其余11对线粒体、叶绿体引物均不能单独区分6个栽培种。在11对叶绿体特异性引物PsbB-PsbT检测多态性,PIC最高,为0.74,两对特异的线粒体SSR标记BnTR 1和BnTR 4中均检测到多态性,多态性条带分别为3和4,PIC值分别为0.5和0.76。
表2 叶绿体与线粒体多态性引物的名称、序列

引物名称细胞器类型 引物序列(F)引物序列(R)ccmp2叶绿体GATCCCGGACGTAATCCTGATCGTACCGAGGGTTCGAATccmp3叶绿体CAGACCAAAAGCTGACATAGGTTTCATTCGGCTCCTTTATtrnL-F叶绿体TCAATTGCACATTCTAGAATTCTAAGCAATTCAATATGGTTATATATTAGAG rpl16叶绿体GGTTCCGTCGTTCCCATCGCCATAATAATTAGATAAATCTGTTCCtrnE-trnT叶绿体AATGGTATGACTAGCTTATAAGGCTTAACAATGAGATGAGGCAATCpsaA-ycf3叶绿体CGGATCTATTATGACATATCCGAAATATGAATACACTAGATTAGGtrnT-rpoC2叶绿体CCTGGCGGTATCAAGATGCCACTGCCATAATGGTACAGAACTATatpB-rbcL叶绿体GAAGGAATAGTCGTTTTCAAGCATAAATAGAGTTCCATTTCGGTrnM-atpE叶绿体CGGCAGGAGTCATTGGTTCAAAGATTTTGTAACTAGCTGACGrbcL-accD叶绿体CTTATATTCATAAGCGAAGAACAATAACAATAGATGAATAGTCAndhB-rps7叶绿体GGGCCGTTATGCTCATTACG TCCTATTCATGGGGATTCCGCh35叶绿体TGGAAAAGGGGAGTTGTCCGGAATATTACTCTCGAACAGACCh39叶绿体CATGAATTAGTAACTGCATCCTCCTATTCATGGGGATTCCGACP2叶绿体GAAAAATGCAAGCACGGTTTTACGATCCGTAGTGGGTTGCACP4叶绿体TACCCGTATTAGGCACTATTTGTAAGACCACGACTGACP35叶绿体ATTGGCTTACTTCTTGCGGGTTTCCGGGATGTTATTACP38叶绿体GGTTCGTTAGCAGGTTTAGTTCCCTAGCAACACTTTACP47叶绿体TGTACTTATGGGAAAGCGCTGGGTTCTTCTACTTCATTTrnK 叶绿体TCAAATGATACATAGTGCGATACA AATAAAGGATTTCTAACCATCTT TrnQ-TrnS 叶绿体AATCCTGGACGTGAAGAATAA AATCCCTCTCTTTCCGTTGA TrnG 叶绿体CCAAAAGCTGACATAGATGTTA TTTCATTCGGCTCCTTTATG Rps2-RpoC2 叶绿体TCTGATAAAAAACGAGCAGTTCT GAGAAGGTTCCATCGGAACAA Ycf3 叶绿体TTGATCTTTACGGTGCTTCCTCTA TCATTACGTGCGACTATCTCC AtpB-RbcL 叶绿体TCTAGGATTTACATATACAACAT CATCATTATTGTATACTCTTTCA PsbB-PsbT叶绿体CCAAAAACTTGGAGATCCAACTAC TTCCATAGATTCGATCGTGGTTTA ACP1叶绿体GAACGACGGGAATTGAACCGGTGGAATTTGCTACCTTTTTACP10叶绿体GTATTAAATCCGAAACTCACTTGACATAAAACTTGGACP20叶绿体CTCAACCGCCATCATACTCGAAACTTAACCCTCTTTACP29叶绿体GGCCATAGGCTGGAAAGTCTGTTTATGCATGGCGAAAAGGACP33叶绿体AGGGATAGTAATAAGAATAGCAGATGTAAGAACGAAAAACP39叶绿体AGACGGGTGAATAGAGTGGTTATGCTTTTCGACGATatp6线粒体GATTTATAGCATCATTCAAGTAAATACAGGAATCAAATAGGGCTGGTGGorf474线粒体GAATCAAATAGGGCTGGTGGCGGAGCTCCCCGGCAGTCAACCAAGAAAGBnTR1线粒体CCGTTAGGGGTATTTAGTAACTCGACATAATGGCAATGTATCGGACTGBnTR2线粒体TGATAACAGTTTCCTCCTAGTTTGCGAACCTGTTAATTAGCACGGAACTABnTR3线粒体GAGTGCTGGCTGTTATAGTATGGTTATAGGAACAGAAAGCTACGCTAACAABnTR4线粒体GAAGTCCGAGGACCTTTAGTACCAGTAAGTTGTAGGTAGGGGCTTCATBnTR5线粒体CTTACAGTCGAGCTCCTTTGTCACTGTAACTCGACCCCTCATCAACTAATBnTR6线粒体AGTATATTGACAGTGCCCCAAGACAGTTACTCGACTGAAAAGGAGAGGT

注:图中M为Maker,A、B分别是引物PsbB-PsbT、BnTR1。图1 线粒体叶绿体引物在39份材料中的扩增结果
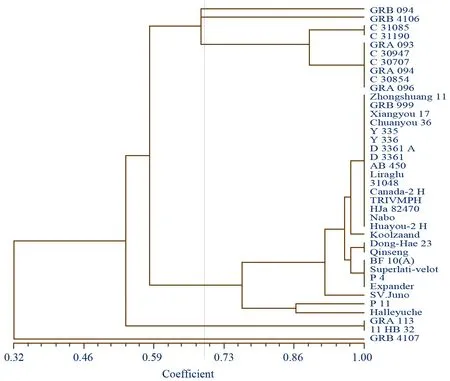
图2 39份材料的遗传聚类结果
2.2 聚类分析
将所有SSR标记结果合并,计算39份材料间的遗传相似系数,采用非加权平均法(UPGMA) 对全部参试材料进行聚类分析,得到的遗传聚类图(图2)。结果显示39份材料的遗传相似系数为0.32~1.00。在遗传相似系数0.71处,39份材料可明显分为6类,与供试材料的分类基本一致,由此表明本研究所设计的标记引物能够有效区分芸薹属6个栽培种的细胞质类型,黑芥与其他5个栽培种遗传距离最远,甘蓝型油菜11 HB 32与甘蓝分为一类。在0.75处甘蓝型油菜分为两大类,来自捷克与南朝鲜为一类,其余25份为一类,0.94~0.98处,甘蓝型油菜具有较丰富多样性。在0.89处芥菜型油菜分为两类,贵州织金与安顺为一类,其余六份为一类。
表3 多态性引物的名称、序列、等位基因数及多态性信息含量

引物名称等位基因长度等位基因数PICccmp2200~30030.24trnL-F200~25020.10rpl16150~20020.47trnE-trnT285~31530.50ACP2100~15030.47ACP4150~20020.15ACP35250~30020.10ACP38250~30020.19ACP47295~31530.48ACP29255~41060.69PsbB-PsbT260~32030.74BnTR1150~20030.50BnTR4230~26040.76
3 讨 论
本研究从不同国家,不同地区200份栽培种筛选出39份具有代表性和遗传多样性芸薹属栽培种,其鉴定引物序列来自于国内外分子育种研究者报道[15-16],具有丰富的多态性,利用31对叶绿体和8对线粒体SSR分别筛选到11和2对稳定且多态性好SSR引物,能较好的区分6个栽培种,这与张瑞杰等[13]的研究结果相似。由于2对引物在鉴别芸薹属中明显呈现5个单倍型[19],在13对线粒体特异性引物中,只有BnTR 1、BnTR 4出现高度多态性。
叶绿体、线粒体基因在甘蓝型油菜与芥菜型油菜均有多样性,表明除了细胞核基因以外细胞质基因组也有多样性,这为芸薹属栽培种分类提供了新方法。目前对油菜品种鉴定均引用核基因组SSR分子标记,尚未利用叶绿体、线粒体分子标记对作物品种进行登记或者认定,本研究的方法可使后续的理论研究和实际生产提供新技术。目前对植物细胞质鉴定未见报道,国内外研究者对芸薹属栽培种相互间作种的远缘杂交,使其后代细胞质基因组较为混乱,出现各种败育类型,部分不育系统核质互作找到恢复源,使得其可利用杂种优势,部分胞质不育出现微粉,可能细胞质基因组相互间排斥或出现变异,或其他可能,这使得鉴定细胞质极为必要,以便更好研究杂种优势在细胞质的分子机理。本研究利用芸薹属6个基本种39份材料,除了27份甘蓝型油菜,芥菜型油菜8份,其他栽培种白菜型油菜1份,甘蓝1份,黑芥1份,埃色俄比亚芥1份,样本量少,未知其他栽培种内遗传多样性,可望后续研究中对其他栽培种进行种内胞质鉴定。
