暗紫贝母FPS基因克隆与表达特性分析
2019-12-16
(内江师范学院生命科学学院/特色农业资源研究与利用四川省高校重点实验室, 四川 内江 641112)
暗紫贝母(FritillariaunibracteataHisao et K.C.Hsia)为百合科多年生草本植物,是川贝母的基原植物,为四川重要的道地药材之一,主产于川西高原地区,具有清热润肺,止咳化痰,镇静平喘,抗炎降压等多种功效[1-2]。已有研究表明,暗紫贝母的有效成分主要为其次生代谢产物生物碱类,此外还包括皂苷、核苷类等,其中异甾体生物碱成分所含比例最多[3]。
对于植物次生代谢途径的研究,其关键酶基因的克隆与分析对于阐明代谢产物合成途径和调控机制至关重要。贝母中异甾体生物碱合成途径与三萜类化合物类似,主要由甲羟戊酸途径合成[4]。法尼基焦磷酸合酶(Fanesyl Pyrophosphate Synthase,FPS)是该途径中的重要限速酶之一[5],其催化2个异戊烯基焦磷酸(IPP)和1个二甲丙烯基焦磷酸(DMAPP)分子发生缩合反应生成法尼基焦磷酸(Farnesyl Pyrophosphate,FPP)[6-7],FPP进一步通过一系列的氧化还原反应合成生物碱等次生代谢产物[8-9]。目前已从腊梅[10]、刺五加[9]、姜花[11]、洋甘菊[12]、虎眼万年青[13]、雷公藤[14]、浙贝母[15]、茅苍术[16]等多种植物中克隆出编码FPS的cDNA序列,但尚未见暗紫贝母异甾体生物碱合成途径中FPS基因的相关研究报道。本研究克隆编码暗紫贝母法尼基焦磷酸合酶的FrFPS基因,对其在不同器官中的表达特性进行分析,并测定不同海拔高度鳞茎的生物碱含量,为研究该基因在暗紫贝母生物碱合成途径中的作用机制提供参考。
1 材料与方法
1.1 材 料
供试暗紫贝母采自四川省阿坝州松潘县,经内江师范学院陈文年教授鉴定为百合科植物暗紫贝母。用清水洗净杂质,滤纸吸干表面水分后的鳞茎作为暗紫贝母总RNA提取材料,液氮速冻保存。表达分析时,在3 200 m、3 400 m、3 600 m、3 800 m 等4个不同海拔高度取材,将暗紫贝母鳞茎、茎、叶分别作为总RNA提取材料,液氮速冻保存。每个海拔高度设3个生物学重复,每个重复分别用5株暗紫贝母相应部位材料混样。
RNAisoPlus试剂,HiScript®Ⅱ 1st Strand cDNA synthesis试剂盒,3′-Full RACE Core Set with PrimeScriptTMRTase,5′-Full RACE kit with TAP,SYBR®Premix Ex TaqTM(Takara 公司);Phanta®Max Super-Fidelity DNA Polymerase(Vazyme Biotech公司);质粒小提试剂盒、琼脂糖凝胶DNA回收试剂盒、pGM-T克隆试剂盒、TOP-10感受态细胞(天根生化科技有限公司),其他试剂为国产分析纯。PCR扩增引物由生工生物工程(上海)股份有限公司合成。
1.2 总RNA提取与反转录
将液氮速冻的暗紫贝母组织迅速研磨成细粉,分装至1.5 mL离心管中,按照RNA提取试剂盒说明书提取暗紫贝母总RNA。用琼脂糖凝胶电泳快速检测RNA的完整性,紫外分光光度计测定纯度和浓度。取500 ng左右的RNA,采用HiScript®Ⅱ1 st Strand cDNA synthesis kit按照说明书反转录cDNA。
1.3 暗紫贝母FrFPS基因的克隆
通过在NCBI上相近物种保守序列的分析,设计1对兼并引物(FPS-DF 1:5′-YCTTGATGATATHATGGACAA-3′;FPS-DR 1:5′-CTYTTGTARATCTTRTGCARG-3′),以暗紫贝母cDNA为模板进行同源片段的扩增。20μL反应体系;94 ℃ 5 min;94 ℃30 s、55 ℃30 s、72 ℃ 2 min,30个循环;72 ℃延伸5 min。得到单一条带后回收PCR产物直接进行正反向测序。
根据保守序列测序结果,按照RACE试剂盒说明书设计3′-RACE引物FPS-3′F 1(5′-GCTTCGGGGCAGATGCTTGATTTGAT-3′)和FPS-3′R 1(5′-ATTTCACTATAGGTTTTTTTTTTTTT-3′)。以暗紫贝母cDNA为模板,反应体系为20 μL,反应程序为:94 ℃ 3 min;94 ℃ 30 s、55 ℃ 30 s、72 ℃ 2 min,30个循环;72 ℃ 5 min。PCR扩增获得基因3′端后进行TA克隆,转化大肠杆菌,挑取阳性转化子测序。
根据保守序列测序结果设计5′-RACE引物FPS-5′F 1(5′-CTGATGATCAGTCGATG-3′)和FPS-5′R 1(5′-CCTCGCCTTGTGTGAGAGTTGTCCAT-3′),以暗紫贝母cDNA为模板,按照RACE试剂盒说明书进行PCR反应获得基因的5′端。电泳后将得到的单一条带进行切胶回收,同时进行TA克隆,选取阳性转化子进行测序。
cDNA序列拼接利用DNAMAN 6.0软件进行,获得cDNA全长,并利用NCBI数据库的ORFfinder查找完整的开放阅读框(ORF)。根据拼接的序列设计包含起始密码子的上游引物FPS-ORF-F(5′-CCATCTATAGCTCCGACGATGGC-3′)和包含终止密码子的下游引物FPS-ORF-R(5′-CTACTTCTGCCTCTTGTAAATC-3′),以cDNA为模板扩增基因的完整开放阅读框。反应体系20μL,反应程序为:94 ℃ 5 min;94 ℃ 30 s、56 ℃ 30 s、72 ℃ 90 s,30个循环;72 ℃ 5 min。PCR产物经TA克隆转化大肠杆菌TOP 10,挑取阳性转化子测序。
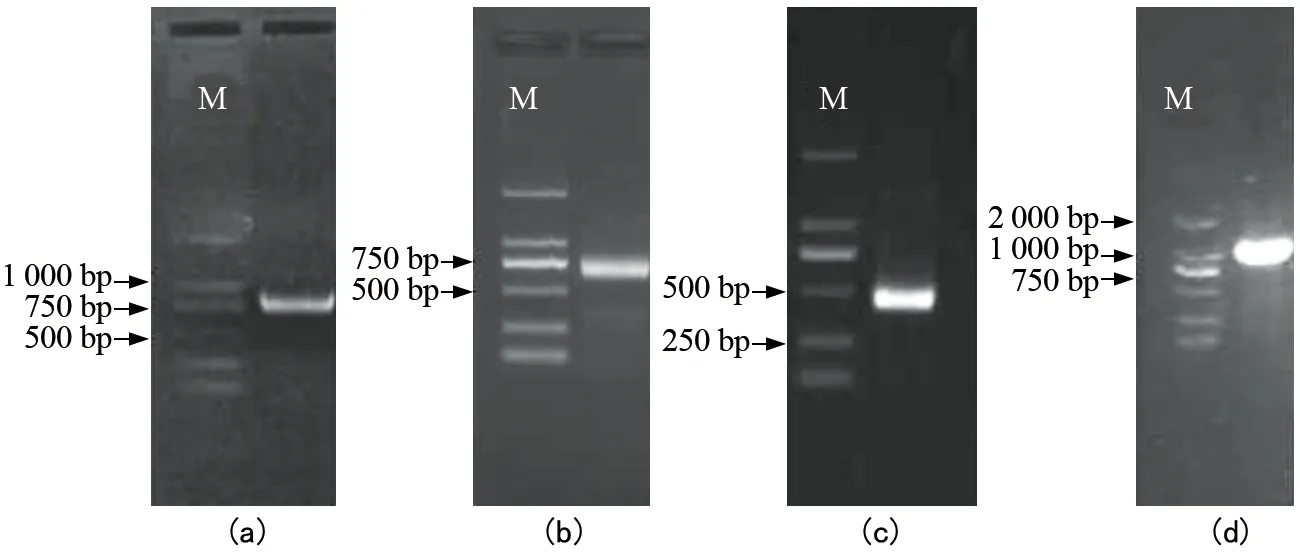
注:(a)为核心片段;(b)为3′-RACE;(c)为5′-RACE;(d)为FrFPS基因的开放阅读框。图1 FrFPS基因克隆电泳结果
1.4 序列的生物信息学分析
利用NCBI的BLAST(http://www.ncbi.nlm.nih.gov./BLAST/)进行基因和蛋白质序列同源性比对。ExPASy的ComputepI/MW在线分析蛋白质的相对分子质量、等电点等理化性质。NCBI在线分析工具CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)分析氨基酸保守结构域。SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_server.html)在线分析蛋白质的二级结构,并通过SWISS-MODEL(https://swissmodel.expasy.org/)进行蛋白质三维结构预测。SignalP 4.1(http://www.cbs.dtu.dk/services/SignalP/)进行信号肽分析。PSORT(https://www.genscript.com/psort.html)预测蛋白质的亚细胞定位。ClustalX 2.1和MEGA 6.0进行序列的多重比对并构建系统发育树。
1.5 FrFPS基因响应不同海拔梯度应答的表达分析
根据克隆的暗紫贝母FrFPS序列,设计荧光定量PCR上游引物FrFPS-F:5′-AGGCGAGGTCAGCCTTGTTG-3′和下游引物FrFPS-R:5′-CATCTGCCCCGAAGCAGTCT-3′,以暗紫贝母18 srRNA(KF 906209.1)为内参,设计上游引物Fr 18 s-F:5′-GCGACGTTCGCTCTCTATCCATAC-3′和下游引物Fr 18 s-R:5′-TGGTTCACGGGATTCTGCAA-3′。引物使用在线Primer 3软件设计(http://frodo.wi.mit.edu/cgi-bin/primer 3/primer 3_www.cgi)。分别提取海拔梯度在3 200 m、3 400 m、3 600 m和3 800 m的暗紫贝母鳞茎、茎、叶的RNA,反转录cDNA作为模板,使用Takara的SYBR®PremixEx TaqTM试剂盒进行qRT-PCR检测,在7300 Realtime PCR System上进行扩增并实时收集数据。15μL反应体系[SYBR Premix Ex Taq(2×)7.5μL;ROX Reference Dye(50×)0.3μL;上下游引物(2μM)各1μL;cDNA模板2μL;ddH2O 3.2μL],反应程序为:95 ℃ 2 min;95 ℃ 15 s,60 ℃ 1 min,40个循环。每处理设3次重复,数据分析得到各个样品的CT值,实验结果采用2-ΔGt的方法进行计算。
1.6 不同海拔高度暗紫贝母生物碱含量测定
采集不同海拔高度(3 200 m、3 400 m、3 600 m和3 800 m)的暗紫贝母鳞茎,洗净烘干过60目筛,经前处理后上柱,分别以贝母甲素、贝母乙素和贝母辛为标准品,HPLC测定鳞茎中的生物碱含量,每个海拔高度3次生物学重复,比较不同海拔高度上鳞茎中生物碱含量的差异。
2 结果与分析
2.1 RACE获取FrFPS基因全长
以暗紫贝母cDNA为模板,根据NCBI数据库中已知的其他物种FPS基因保守序列设计简并引物,PCR扩增获得732 bp的核心序列。根据获得的中间片段,分别设计引物进行3′-RACE和5′-RACE,PCR扩增产物经测序表明获得基因的3′和5′端序列,分别为913 bp和498 bp。利用DNAMAN 6.0软件将5′和3′端序列进行拼接,获得1条具有完整开放阅读框(ORF)全长为1 333 bp的序列。设计包含起始密码子和终止密码子的特异引物扩增基因的ORF,验证获得的开放阅读框序列长度为1 059 bp,编码352个氨基酸(图1、图2)。将该基因命名为FrFPS,序列已提交到NCBI,获得的登录号为KX 900485。
2.2 暗紫贝母FrFPS基因的生物信息学分析
ExPASy在线预测FrFPS基因的编码蛋白质分子量为40.13 KDa,等电点5.17。通过在NCBI网站中进行同源性比对,发现其编码蛋白质与其他植物法尼基焦磷酸合酶氨基酸序列具有较高的同源性,其中与同为百合科植物的铁炮百合氨基酸序列同源性最高,达到95%,其次为同科植物海葱83%,与油棕82%,与竹节参、蕙兰、喜树、菠萝、腊梅等的相似性均为81%。相似性在75%以上的植物达数十种,表明植物法尼基焦磷酸合酶保守性较高。
SOPMA在线分析表明,FrFPS基因编码的蛋白质二级结构以α-螺旋为主,占60.51%;其次是无规则卷曲,占21.31%;此外还包括9.94%的延伸链和8.24%的β-转角(图3)。利用NCBI在线分析工具CDD分析保守区,发现FrFPS的功能域氨基酸具有5个保守结构域,属于Isoprenoid-Biosyn-C 1蛋白家族。通过ClustalX 2.1序列比对表明,FrFPS保守结构域氨基酸组成与其他植物一致,其中有2个高度保守的富含天冬氨酸区域(DDXXD)(图4),说明各物种FPS基因在进化过程中相对保守。进一步利用SWISS-MODEL构建该蛋白质的三维结构模型,结果显示FrFPS蛋白主要由多个α-螺旋和无规则卷曲组成,α-螺旋在表面和内部分布较均匀(图5)。PSORT预测分析显示,FrFPS编码蛋白质定位于细胞质、线粒体基质空间及微体上。SignalP 4.1分析表明FrFPS蛋白中不存在信号肽,为非分泌蛋白。

图2 FrFPS基因全长cDNA序列及其推导的氨基酸序列

图3 FrFPS推导蛋白二级结构预测图
为确立FPS基因在不同植物间的进化关系,将FrFPS基因编码的氨基酸序列与GenBank数据库中登录的铁炮百合、青蒿、紫花苜蓿、黄芪、烟草、小麦等21个物种的法尼基焦磷酸合酶氨基酸序列进行比对分析,基于邻接法(NJ法)构建系统进化树(图6)。结果表明,包含FrFPS蛋白的共22种氨基酸序列可以聚为两大类。第一类中,暗紫贝母FPS与同为百合科的铁炮百合(Liliumlongiflorum,JF 273657.1)和海葱(Ornithogalumlongebracteatum,KF 509889.1)聚为一个小亚类,然后与芭蕉科的小果野蕉(Musaacuminata,XM_009396239.2),棕榈科的海枣(Phoenixdactylifera,XM_008794576.2)、油棕(Elaeisguineensis,XM_010925807.2),姜科的姜花(Hedychiumcoccineum,AER 12202.1),凤梨科的菠萝(Ananascomosus,XM_020250595.1),禾本科的小麦(Triticumaestivum,JX 235714.1)和穇子(Eleusinecoracana,KT 824871.1),木兰科的乐昌含笑(Micheliachapensis,GQ 214406.1)依次聚合在一起,共同构成一个大的分枝。说明暗紫贝母在进化上与这些植物的亲缘关系较近。第二类包括豆科的黄芪(Astragalusmembranaceus,KP 233831.1)、绿豆(Vignaradiatavar.Radiata,XP_014517551.1)、紫花苜蓿(Medicagosativa,ADC 32809.1),茄科的烟草(Nicotianatabacum,XM_016600471.1),蔷薇科的玫瑰(Rosarugosa,KP 768082.1),菊科的青蒿(Artemisiaannua,U 36376.1)等,说明暗紫贝母在进化上与这些植物亲缘关系较远。

图4 FrFPS序列与其他FPS氨基酸序列多重比对
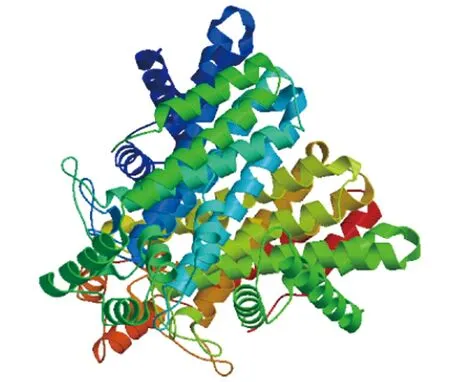
图5 FrFPS三级结构预测图
2.3 暗紫贝母FrFPS基因响应海拔梯度的应答
为研究FrFPS基因表达对不同海拔高度的响应,采用实时荧光定量PCR分析FrFPS基因在3 200 m、3 400 m、3 600 m和3 800 m不同海拔高度上暗紫贝母鳞茎、茎和叶中的表达情况。结果表明,FrFPS基因主要在暗紫贝母鳞茎中表达,叶片和茎中表达量极低。在不同海拔梯度上,FrFPS基因表达量呈现出随海拔梯度升高而降低的趋势。较低海拔(3 200 m、3 400 m)暗紫贝母鳞茎中FrFPS基因表达量约为高海拔(3 800 m)鳞茎表达量的6倍左右(图7)。表明较低海拔更有利于FrFPS基因的表达。
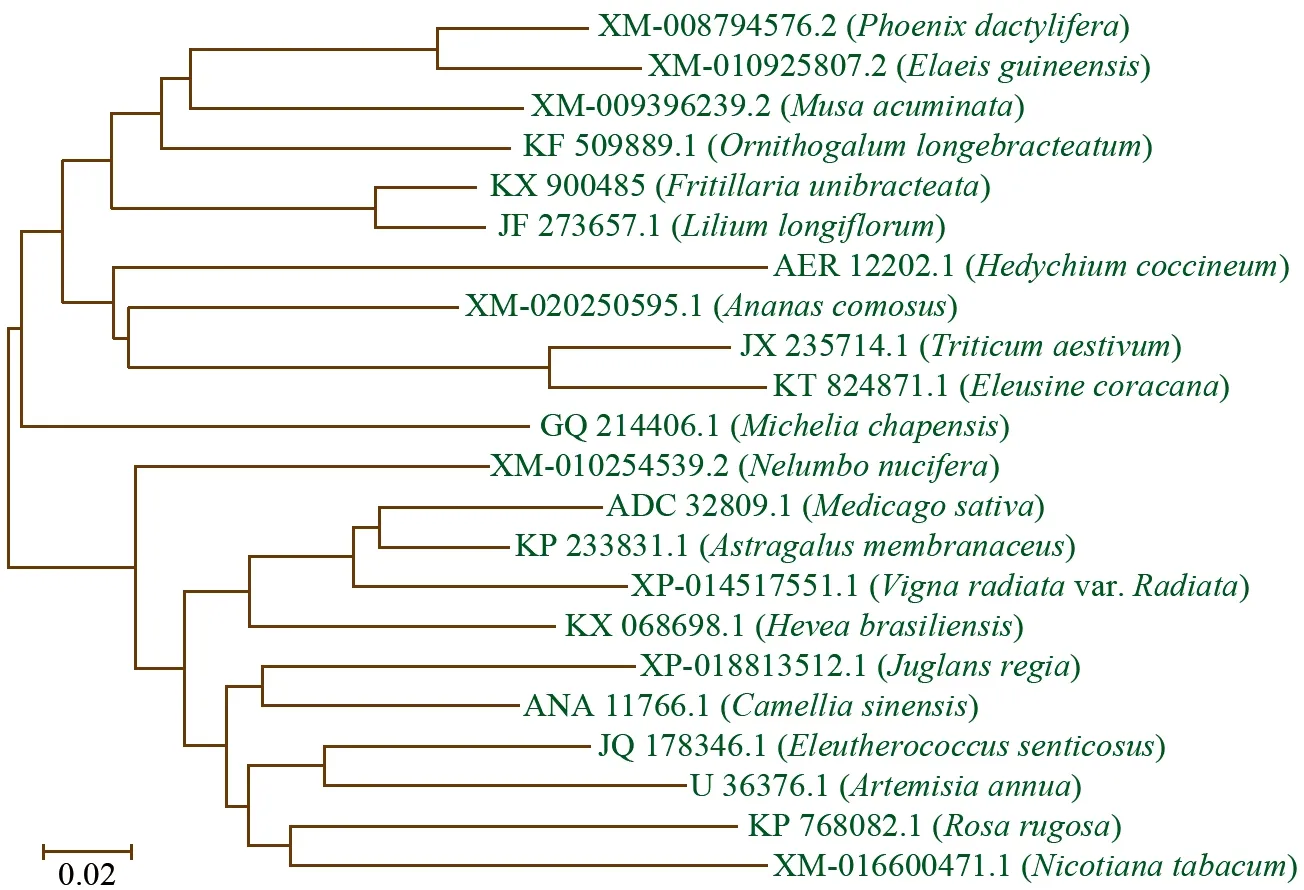
图6 FPS蛋白系统进化分析
2.4 不同海拔高度暗紫贝母生物碱含量
HPLC测定暗紫贝母鳞茎生物碱含量结果表明,不论以贝母辛或贝母甲为标准品,较低海拔高度(3 200 m和3 400 m)的暗紫贝母鳞茎中的生物碱含量要高于较高海拔(3 600 m和3 800 m)的鳞茎,以海拔3 200 m鳞茎中生物碱含量最高(表1)。这与实时荧光定量PCR测定结果相似。在鳞茎中未能检测出贝母乙素。
表1 不同海拔高度暗紫贝母鳞茎生物碱含量

海拔高度(m)生物碱含量/(μg·g-1)(以贝母辛为标准品)含量/(μg·g-1)(以贝母素甲为标准品)320062.9618.23340047.8813.86360047.3713.71380046.0213.32

图7 FrFPS基因在不同海拔高度的表达分析
3 讨 论
法尼基焦磷酸合酶(FPS)是一种异戊烯基转移酶,是类异戊二烯途径的一个重要调节酶,催化异戊烯基焦磷酸(IPP)和二甲基丙烯基焦磷酸(DMAPP)合成法尼基焦磷酸(FPP)。FPP是植物体内甾体、皂苷、倍半萜等多种次生代谢产物的前体,进一步通过鲨烯合酶催化甾体类、皂苷类、倍半萜类等物质的合成[8,17-18]。
本研究克隆的FrFPS基因的编码产物具有植物法尼基焦磷酸合酶的一般特征:二级结构以α-螺旋为主,其次是无规则卷曲,β-折叠和β-转角较少,分散于整个蛋白质中[18]。以IPP为底物的异戊烯基转移酶等,均含有典型的DDXXD保守结构域[17]。本研究中的暗紫贝母FrFPS基因所推测的肽链的103~107氨基酸和242~246氨基酸处同样有典型的DDXXD保守结构域(图4),这与茅苍术[16]、三七[17]、陆地棉[18]等植物的FPS基因相似,2个区域富含天门冬氨酸,认为是这类酶的活性中心[17,19]。FrFPS跨膜区及亚细胞定位分析表明其无跨膜结构,定位于细胞质,这符合植物FPS不存在跨膜结构域的观点。通过进化树分析,暗紫贝母与百合科植物铁炮百合亲缘关系较近,体现了其在分子系统演化关系中所处的位置,为其分类地位提供了佐证,也为进一步认识植物基因功能提供依据。
已有研究表明,植物FPS表达具有组织特异性[20-22],并且伴随着类异戊二烯衍生物含量而增加[23]。本研究中FrFPS在暗紫贝母鳞茎中表达量最高,茎、叶中虽有表达但量非常低,说明FrFPS具有组织特异性,同时也符合暗紫贝母以鳞茎入药的传统。本课题组在前期研究发现,暗紫贝母各物候期都是在低海拔部位最先出现,在高海拔部位最迟出现;鳞茎平均单粒重量有随海拔升高而降低的趋势[24]。本研究中,较低海拔(3 200 m、3 400 m)暗紫贝母鳞茎中FrFPS表达量为高海拔(3 800 m)鳞茎中的6倍左右,FrFPS表达可能是暗紫贝母鳞茎在不同海拔梯度上存在差异的原因之一,也为暗紫贝母人工栽培的适生海拔选择提供了分子水平的参考依据。但暗紫贝母的生长发育及次生代谢产物合成受多种环境因子的影响,对其他代谢过程中的关键酶基因作用及代谢调控机制还需进一步研究。
利用FPS基因过量表达可以有效提高青蒿[25]、人参[26]等药用植物活性成分的产量,在药用植物遗传改良中具有重要作用。本研究首次克隆获得暗紫贝母FPS基因的全长cDNA,并对其响应海拔高度的表达特性进行了分析,为进一步探索暗紫贝母法尼基焦磷酸合酶在生物碱合成途径中的作用机制奠定了基础。
