菌根植物适应低磷胁迫的分子机制
2019-12-13郝志鹏陈保冬
李 芳,郝志鹏,陈保冬*
(1 中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085;2 中国科学院大学,北京 100049)
磷 (P) 是植物体内很多重要有机化合物如核酸、磷脂等的组成成分[1],参与植物体内的许多基础代谢过程,如氮素代谢、脂肪代谢等,同时也是调节能量代谢、酶促反应和信号转导过程所必需的元素[2]。在酸性土壤中,磷酸盐与铝和铁形成难溶复合物,而在碱性土壤中磷酸盐与钙络合,导致土壤溶液中磷酸盐浓度极低,造成土壤对作物供磷不足[3]。在农业生产中,磷肥施入土壤后大约有85%~90%变成植物难以利用的形态[4],施肥当季磷利用率普遍很低,低磷胁迫成为制约农作物生长的重要因素[5]。另一方面,磷肥的主要生产原料磷矿粉是不可再生的自然资源,但为了确保作物的生产力,施用磷肥已经成为常态,无论从经济角度或者生态角度考虑这种做法都是难以持续的,因此提高土壤磷的利用效率一直以来都是土壤和植物营养学家的一个挑战。
植物长期生长在低磷环境中,进化出一系列适应机制,如增加根冠比与侧根长度、释放有机酸活化土壤磷等。植物与菌根真菌形成共生体系也是植物对低磷胁迫的一种重要适应机制[6]。菌根是指土壤菌根真菌侵染植物营养根所形成的共生体系。丛枝菌根真菌 (arbuscular mycorrhizal fungi,AMF) 是菌根真菌的重要类群,遍布于所有陆地生态系统,能够与绝大多数陆生高等植物形成共生关系,其生物量约占土壤微生物总量的5%~50%[7-8]。AMF 能改善宿主植物的矿质营养和抗逆性,特别是可以有效增强宿主磷吸收能力[9]。AMF 能扩大宿主植物根系的吸收面积[10],释放有机酸和土壤酶活化土壤磷[11]。同时,AMF 磷酸盐转运蛋白与磷酸盐的亲和力高,菌丝中磷的转运速率比根中转运快[12]。应用AMF 能够大幅度提高作物对土壤磷的利用效率,减少磷肥施用,有利于农业可持续发展[13]。
目前已有很多研究从不同角度揭示了菌根植物适应低磷胁迫的生理机制,近年来新的分子生物学技术如高通量测序、转录组学及代谢组学的应用,使得我们对宿主植物和菌根真菌协同适应低磷胁迫的分子和信号机制有了更为深入的认识,但这些研究还比较分散,缺乏系统性的总结。菌根共生体系感知和适应磷缺乏的分子调控网络仍旧存在很多研究空白。基于此,本文综述了国内外关于菌根植物适应低磷胁迫分子响应机制方面的研究进展,分析当前研究工作中存在的不足,并提出未来应当加强的研究。
1 植物和菌根真菌感知磷胁迫信号
磷不仅作为营养物质被菌根植物吸收利用,同时又可以作为一种信号来调控共生体系的建成和发育。细胞内磷浓度的增加可以抑制磷饥饿反应,因此也可以将磷看作信号物质[14],亚磷酸盐或磷酸酯(Phi) 是磷酸盐类似物,能被植物磷酸盐转运蛋白吸收,但是在细胞内不能被氧化为磷酸盐 (Pi) 而参与代谢。研究发现在低Pi 条件下,外源施加Phi 减弱甚至中止磷饥饿反应[15],Phi 和Pi 之间结构相似并且Phi 不能作为营养物质,因此Phi 是作为信号调控植物磷饥饿反应的。研究表明,土壤中磷浓度的变化可以影响植物根系的形态结构[15]。总而言之,磷既作为营养元素又作为信号物质调控植物生长发育。那么,植物与AMF 是怎么感受磷信号的呢?
以前的研究认为根细胞膜定位传感器和细胞内传感器分别感知外部磷酸盐浓度和内部磷营养状态的变化[16]。植物的生长是由内源性信号和环境因子共同调节的,而植物对于磷信号的感知也取决于两个方面,其一是对环境介质中的磷浓度的局部感知[17-19],二是植株内部的磷状态调节[20-21]。局部Pi 浓度传感和信号传导可以启动根系结构调整以增强根系对Pi 的摄取能力,无论枝条中的磷状态如何,初级根尖与含磷基质的物理接触是必要的,这使得局部Pi 信号被植株感知[26],而植物本身磷营养状态或长距离信号传导则用于调节植株体内Pi 的活化和再分配[22-23]。基于分根培养实验的转录组学分析显示,参与Pi 摄取重获和脂质代谢的基因通常是系统性调节的,而与应激或激素反应相关的基因在局部受到调节[19]。土壤中的Pi 主要由植物根系吸收,通过磷酸盐转运蛋白(phosphate transporter,PT) 转运到植物体内[24]。在很多情况下,尚不清楚实际参与Pi 转运的转运蛋白的详细信息。磷信号被相应功能的磷转运蛋白感知从而传递到植物体内,植物体内存在两种形式的磷素运载模式,一是低磷条件下对磷具有高亲和力的高效吸收转运模式,二是对磷亲和力低的组成型运载模式[25],根系会根据感受到的不同磷信号产生不同的响应,如磷的转运、根系构型改变、根分泌物变化、代谢通路改变、脂质重塑与促进菌根共生等。有研究发现植物内部的感受器如P D R 2(P 5 型ATP 酶)、LPR1/LPR2 等能够感受外部磷信号的变化[26],调节分生组织的活性,在磷缺乏条件下植物根分生组织耗竭而使根系变得短粗[27]。对于植物养分吸收来说,外界磷浓度的变化会引发植株不同的反应,然而对于磷信号的定量研究仍旧非常匮乏,磷作为营养元素和信号物质,目前尚不能明确哪一“身份”在后续的植物生理反应中起主导作用[14]。
菌根植物通常有两条磷吸收途径:直接通过根表皮和根毛 (直接途径) 与间接通过AMF 根外菌丝吸收进而转运到根皮质细胞 (菌根途径)。在根外菌丝中,Pi 分别由H+-ATP 酶和Na+-ATP 酶驱动通过H+/Pi 共转运系统和Na+/Pi 共转运系统摄取,并通过液泡转运蛋白聚合物 (VTC) 在液泡中聚合成多聚磷酸盐 (Poly-P)[28]。将AMF 从磷缺乏的培养基转到磷充足培养基中培养时,Pi 会快速聚集并且形成大量的Poly-P[29]。目前对于Pi 是如何通过菌丝体与根细胞之间的质外体界面仍旧未知[30]。Poly-P 可能经由AMF 水通道蛋白 (AQP 3) 介导的水流长距离运输到宿主中[31-32](图1)。Pi 通过液泡多磷酸酶从Poly-P 释放,通过液泡Pi 输出器 (PHO91) 输出到胞质溶胶中,然后掺入ATP 中进行聚合。通过这些过程,AMF 能够在数小时内积累大量Poly-P 而不会对细胞Pi 水平产生扰动,使得AMF 细胞Pi 稳态得到严格调节[33]。
在缺磷条件下,菌根真菌与植物根系更容易建立共生体系[34]。Pi 缺乏刺激根中独脚金内酯 (SL) 生物合成并分泌到土壤中。SL 作为根际信号调节宿主植物与AMF 的相互作用,诱导AMF 分支和菌根侵染[35]。反之,高水平的磷营养供给会抑制共生体系的形成。高磷浓度下,类胡萝卜素 (SL 的前体) 生物合成相关基因的表达受到抑制,菌根特异性诱导的PT 基因表达下调[36],同时植物的ABC 转运子PDR1基因的表达受到抑制。PDR1 调控SL 向细胞外的分泌,在调节AMF 发育过程中起到关键作用[37],PDR1基因表达受抑最终会导致高磷胁迫下菌根侵染率下降[38]。不同AMF 种类对于磷浓度的响应表现出差异[39],不同的磷信号也会引发植物和AMF中不同的生物过程响应[40],例如在不同磷水平下菌根植物中与磷饥饿相关的转录物质的积累表现出很大差异。
2 菌根植物适应低磷胁迫的分子机制
2.1 调控植物激素及有机酸、磷酸酶合成相关基因的表达
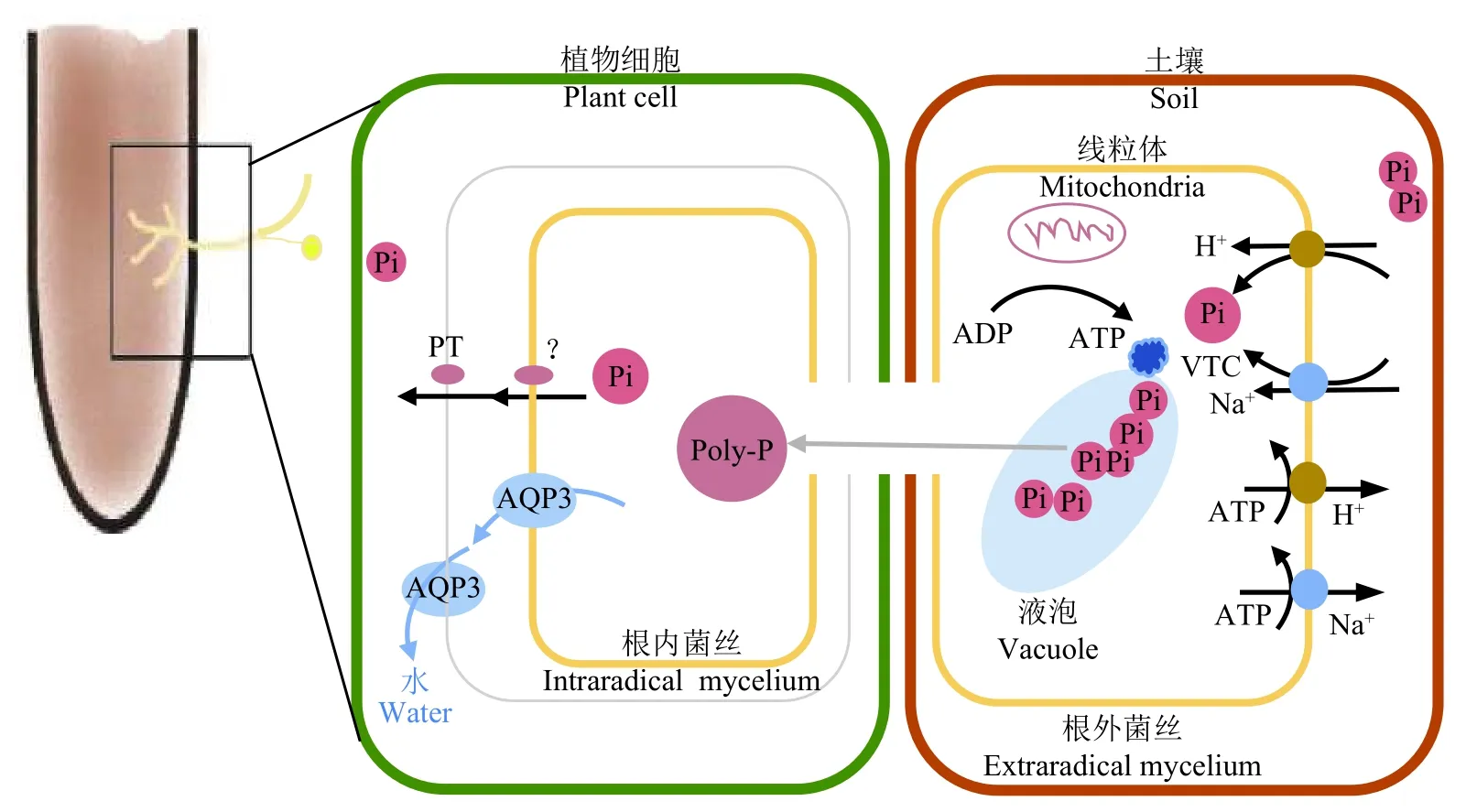
图 1 丛枝菌根共生体系中Pi 吸收与转运示意图Fig. 1 Schematic diagram of Pi uptake and translocation in arbuscular mycorrhizal symbiosis
植物激素是植物发育和应对环境胁迫的关键调节剂[41],通过影响根系生长和糖信号传导来调节植物对磷的摄取。应用基因芯片技术发现了在磷胁迫下激素生物合成基因在转录水平的变化[42]。有充分证据表明,在缺磷条件下植物根系结构显示出很大的可塑性,生长素、乙烯、细胞分裂素、独脚金内酯、赤霉素和脱落酸等都参与了对根系结构的调控[14]。如参与乙烯合成第二步的1-氨基环丙烷-1-羧酸氧化酶(ACO) 基因在Pi 缺乏下上调[43],而乙烯参与Pi 缺乏诱导的根系结构重塑,可以抑制初生根生长,促进侧根生长,以此扩大根系与土壤的接触面积,提高植物的磷摄取能力[44]。
菌根化和施加磷肥都能提高植物细胞分裂素的含量,从而改善植物生长。菌根侵染也会特异性诱导脱落酸生物合成增加[45]。众所周知,脱落酸、茉莉酸和水杨酸能够增强植物对生物和非生物胁迫的耐受性[46]。很多研究中发现,低磷胁迫下参与脱落酸和茉莉酸生物合成和信号传导的基因在AM 植物叶中的特异性上调。例如,在接种AMF 的玉米中,参与茉莉酸生物合成的基因 (如编码12-oxophytodienoate reductase OPRs、3-hydroxybutyryl dehydrogenase protein 的基因) 表达上调。前文提到的独脚金内酯也是一种植物激素,低磷环境会刺激植物分泌独脚金内酯,促进菌根共生体系建立从而缓解低磷胁迫。
有机酸 (如柠檬酸、苹果酸和草酸) 可以将Al3+、Fe3+和Ca2+结合的Pi 释放出来,从而提高土壤中有效磷含量。在磷缺乏土壤中很多植物有机酸合成与分泌相关基因会被高度诱导[47]。研究发现AMF 也能够分泌有机酸和磷酸酶活化土壤磷,低磷条件下被AMF 侵染的万寿菊根中特异性的磷酸酶活性高于供磷情形[48]。某些紫色磷酸酶基因在低磷胁迫下参与植物与AMF 共生的反应,两个紫色磷酸酶基因表达在菌根大豆根中显著增加[49]。以枳实生苗为试验材料的盆栽实验,基质中草酸、苹果酸和柠檬酸含量均与菌根侵染率成显著正相关,低磷条件下菌根植物有机酸分泌量明显增加[50],在菌根植物叶片内也检测到有机酸的大量累积[51]。
2.2 调节磷转运蛋白基因差异表达
已知的磷酸盐转运蛋白基因可以分为四大类,分别在植物的不同生理部位发挥作用。PHT1 家族属于高亲和力磷转运蛋白,主要在根中表达[52]。PHT1家族的转运蛋白对于从土壤中摄取Pi 以及从根到茎磷的转运是非常关键的,如耐低磷小麦品种根中TaPHT1;2 的转录水平比低磷敏感小麦品种的表达要高[53]。PHT2、PHT3 和 PHT4 家族还有待探究,其中PHT3 主要在线粒体中,PHT4 在高尔基体中。
研究者们把丛枝菌根涉及的磷酸盐转运蛋白基因分为三种类型:1) 植物的磷酸盐转运蛋白基因;2) AMF 的磷酸盐转运蛋白基因;3) AMF 诱导植物产生的磷酸盐转运蛋白基因[36,54]。AMF 通过调控植物与其本身的碳磷代谢基因,可以促进共生体建成,进而调节宿主植物的磷营养。Sawers 等[55]在30 个不同品种的玉米接种实验中发现,接种AMF上调了大多数磷转运蛋白基因的表达,促进了玉米对磷的吸收,而抑制磷饥饿诱导表达的磷转运蛋白基因的表达,菌根玉米的磷吸收效率与编码PHT1转录物的积累相关。在低磷条件下,接种AMF 或在菌根分泌物作用时均显著上调了玉米根中磷转运蛋白基因Pht1;2、Pht1;6 和G3PT 的表达水平;与高磷情形相比,低磷条件下磷转运蛋白基因与碳磷代谢基因如GiPT、HXK1b、CHS1 和NAG1 表达水平提高了2~3 倍[56]。也有研究报道AM 接种显著提高了AM 特异诱导的磷转运蛋白基因Pht1;6 的表达水平,并且降低了Pi 饥饿诱导型Pi 转运蛋白基因Pht1;3 在玉米根中的表达水平[54],表明AMF 接种缓解了宿主植物的磷饥饿状态,菌根植物以此方式适应磷胁迫。
分别来自Glomus versiforme、Glomus intraradices和Glomus mosseae 的GvPT、GiPT 和GmosPT,是AMF 自身的磷酸盐转运蛋白。除了GmosPT 也在根内菌丝中显著表达,这三种磷转运蛋白都在根外菌丝中表达。转录组数据分析显示,AMF 中除了存在GiPT 这类高亲和力的磷转运系统,还发现了低亲和力Pi 转运体系,还有许多磷酸酶及调节因子也在AMF 中被发现。这表明AMF 中存在与酵母细胞中一样的PHO 调控系统,可以有效调节磷的吸收与转运[57]。AMF 根外菌丝中的磷酸盐转运蛋白从土壤环境中摄取Pi,通过VTC 在液泡中聚合成Poly-P 转运到根内菌丝,然后通过一种未知途径将其转运至植物与菌根真菌之间的“环丛枝空间 (periarbuscular space)”,然后被植物根细胞膜上的磷酸盐转运蛋白转运,从而使得AMF 吸收的磷转运到宿主植物中。AMF 是专性共生真菌,需要与植物形成共生体才能完成其完整的生活史。GigmPT 的敲除降低了AMF 对宿主植物的定殖水平,表明GigmPT 介导的磷酸盐转运对于AM 共生是非常关键的,GigmPT 可能作为磷信号受体在磷酸盐摄取和感应磷酸盐浓度变化方面发挥重要作用[58]。
对于菌根植物来说,除了植物自身的磷转运蛋白基因能够对磷胁迫做出响应,AMF 特异性诱导的磷转运蛋白基因对于植物适应磷胁迫也具有重要作用。目前已从很多植物如番茄[59]、水稻[60]、苜蓿[61]和玉米[62-63]中鉴别出菌根特异性诱导的磷转运蛋白基因,这些基因编码的蛋白是磷酸盐通过质膜转运的重要通道,也是菌根植物适应低磷胁迫的一种重要机制。紫花苜蓿中的MtPT4 是受AMF 特异诱导的一种磷转运蛋白,通过RNAi 技术使MtPT4 基因表达沉默,植株生物量即显著降低,即使接种AMF 对植物生物量也没有显著影响。不仅如此,MtPT4 功能的丧失还影响到AMF 的形态与生长,MtPT4 基因突变的植株中菌根侵染率和根外菌丝密度均显著低于野生型[36]。因此,特定的磷酸盐转运蛋白基因对于菌根共生体来说是不可或缺的。对敲除了菌根特异磷酸盐转运蛋白基因Pht1;6 的玉米进行RNA 测序发现,Pht1;6 缺失导致菌根中激素 (赤霉素、脱落酸与乙烯)、信号传导 (受体激酶、脂肪酸) 和细胞结构(细胞壁、脂质) 等代谢与生长过程失调,这也表明磷酸盐转运蛋白基因具有重要的生物学意义,可能与其他途径共同调节植物生长发育[64]。
2.3 转录因子和miRNA 对低磷胁迫的响应
转录因子在植物生长发育的各个阶段以及植物适应各种逆境胁迫过程中都发挥着重要作用。研究表明转录因子可以通过激活下游靶基因来改变植物对磷胁迫的耐受性[65],而且转录因子在AM 共生体系的发育中也具有重要作用[66]。MicroRNA (miRNA)可以作为长距离信号参与植物低磷胁迫响应,进而参与植物发育调控和逆境响应等重要过程。PHR1(phosphate starvation response 1) 是一种MYB(myeloblastosis-related proteins) 型转录因子[67],在PHR1 过表达的植株中 (玉米、水稻和小麦的PHR1同源序列分别为ZmPHR1、OsPHR2 与TaPHR1-A1)观察到根伸长率的增加、根毛生长以及高亲和力磷酸盐转运蛋白基因表达的上调[68-70],通过这种方式植株提高了磷酸盐的吸收和利用效率以适应磷胁迫。转录因子在菌根共生体的建成与发育中起重要作用[71],菌根转录因子结合序列MYCS 和P1BS 介导了AM 调节的磷酸盐转运蛋白基因的转录激活[72]。有研究发现,一种转录因子MYB1 介导AMF 的丛枝退化,丛枝的退化时间由植物和丛枝的营养状况控制,磷酸转运蛋白PT4 突变体的丛枝退化加速[36],可能是由于缺乏丛枝传递的磷酸盐[73-74]。换个角度考虑,低磷胁迫下菌根植物的丛枝传递的磷营养达不到宿主植物要求,或许会诱导相关转录因子的表达从而介导丛枝退化,不利于菌根共生体发育。然而,有关菌根植物转录因子对于低磷胁迫的特异性响应的研究极少,在今后的研究中应得到重视。
受 PHR1 调控的基因包括PHT1 家族基因、PHO1、At4 和miRNA399 (miR399) 等。研究发现miR399 受缺磷诱导表达,能直接作用于PHO2 (泛素E2 连接酶,是miR399 的靶基因) 基因的 5’-UTR 区,降解PHO2 基因的mRNA,使其表达量下调,从而调控植物对磷的吸收、转运和再利用过程;miR399 的成熟片段在菌根化苜蓿和烟草的根中积累,从而抑制PHO2 表达,促进磷吸收和转运,但是几个原始转录本却仅仅在菌根植物的地上部累积,表明其对低磷胁迫的响应在地上部先于根部。比较有趣的是一个推定的磷酸盐转运蛋白基因在菌根中表达显著上调,而它也是miR399 的靶基因[75-76]。miR5229 的丰度在根皮层中含有AMF 丛枝的细胞中显著增加,其参与了丛枝的特定发育阶段,有利于菌根共生体系的建立,提高植物对低磷胁迫的适应性[76]。
3 总结与展望
AMF 与大多数陆生植物建立的菌根共生体系对于植物磷营养具有重要作用。本文主要从有机酸、磷酸酶与激素、磷转运蛋白基因、转录因子与小分子物质miRNA 等方面讨论了菌根植物对磷胁迫的响应与适应机制 (图2)。环境Pi 浓度作为营养信号诱发菌根植物的生理响应过程,有机酸活化土壤磷,激素在调节基因表达和改变根系构型方面起着至关重要的作用,磷酸盐转运蛋白基因灵敏而高效的表达以应对不同的磷信号,转录因子作为感知信号和调控转录表达水平的枢纽,miR399 充当长距离信号调节植物自身反应。这些因素既有单独作用又相互交叉关联,共同构成菌根植物适应磷胁迫的分子调控网络。磷胁迫信号是一个牵动点,起到提纲挈领的作用,菌根植物低磷胁迫的适应调节机制离不开对磷信号的感知。
目前关于菌根植物适应低磷胁迫机理的研究已有很多,但仍有很多未知领域尚待探索。笔者认为以下几个研究方向应当得到重视。
1) 菌根共生界面的磷转运机制。AM 共生体系对磷的吸收转运过程大部分已经明确,但是根内菌丝中的Pi 是通过何种载体蛋白运输到环丛枝空间从而被宿主植物吸收仍不清楚。低磷胁迫下菌根植物的转录组学研究发现了很多假定蛋白,其中是否有一些蛋白参与了根内菌丝与植物间的磷转运?随着组学技术和大数据分析方法的进步,结合转录组学与蛋白质组学及RNA 干扰等技术,定位根内菌丝的磷运输途径将成为可能,这对于更全面地认识磷在真菌和植物间的转运及调控机制具有重要意义。
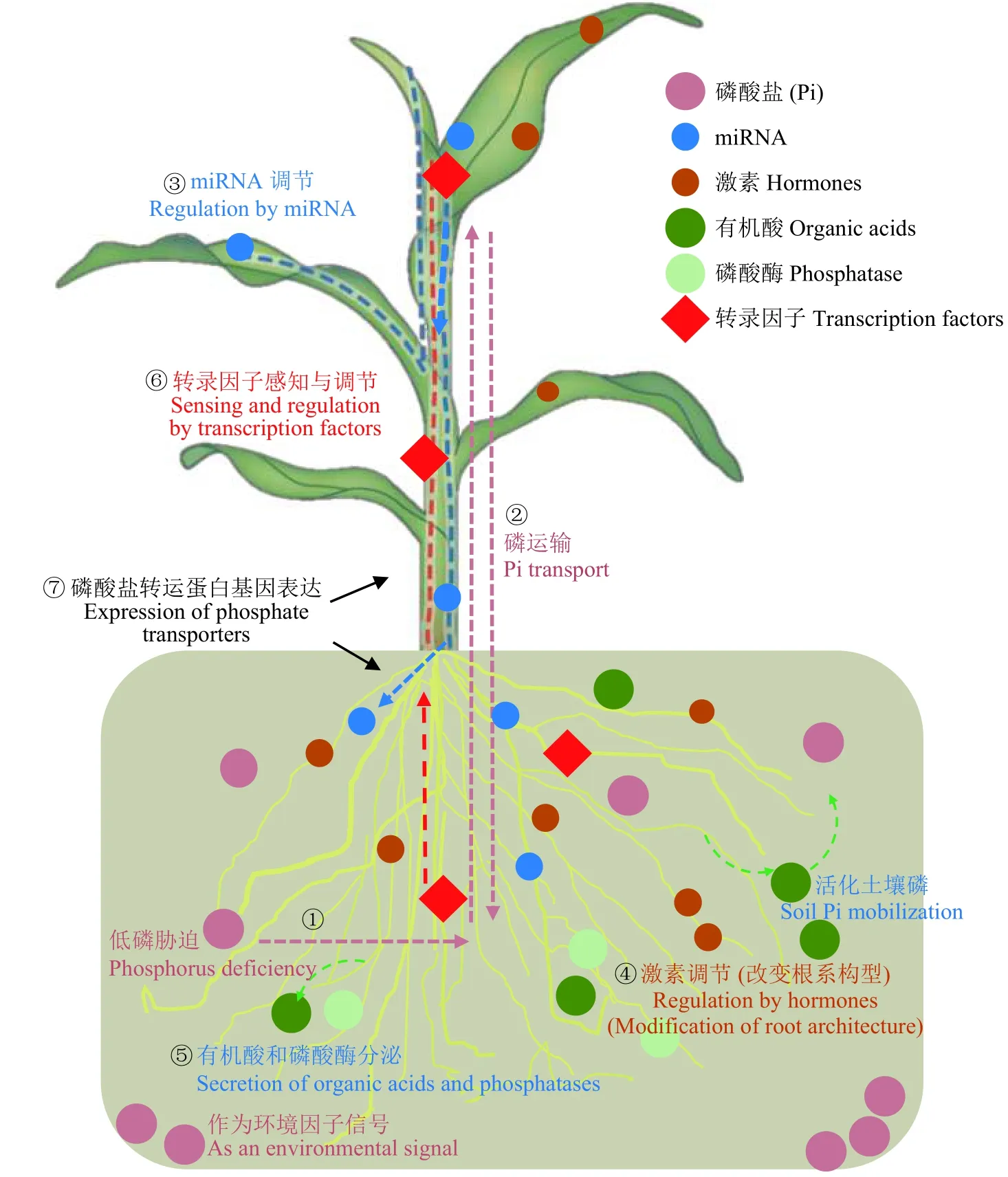
图 2 菌根植物适应低磷胁迫分子机制示意图Fig. 2 Molecular mechanism for mycorrhizal plant adaptation to low P stress
2) 菌根植物适应低磷胁迫的转录因子调节。转录因子对真核生物转录起始过程十分重要,但是有关磷胁迫下转录因子及其作用机制的研究还处于起步阶段,如已知MYB、NAC (NAM-ATAF-CUC2)、bHLH (basic helix-loop-helix) 等转录因子对于植物低磷耐受性具有重要作用,但其各个家族的功能却不能明确区分。菌根植物响应磷胁迫包含了植物本身和AMF 对植物的特异性调控,而AMF 特异性调节宿主植物适应磷胁迫的转录因子有待进一步探究。随着基因组学和转录组学的不断发展,对菌根植物适应低磷胁迫的转录因子调节机制将会有更多新的认识。
3) 各调控因子相互作用。菌根植物适应低磷胁迫的各类调节因子不是独立作用的,如miRNA 通过调节PHO2 进而调控磷转运蛋白基因表达以应对低磷胁迫。目前对菌根植物体内磷信号传导和磷代谢途径交互网络的认识还远远不够,今后的研究要利用新的分子生物学技术,深入挖掘菌根植物适应低磷胁迫的分子调控网络和关键调控因子间的相互作用,从而更全面地揭示植物和菌根真菌协同适应低磷胁迫的分子机制,为发展和应用菌根技术调控植物磷营养奠定理论基础。
