miR-509-3p靶向BCL2基因调控乳腺癌细胞增殖、侵袭、迁移及凋亡的分子机制
2019-12-10秦少杰刘清媛唐振宁刘奇伦
秦少杰 刘清媛 唐振宁 刘奇伦
(宁夏医科大学总医院肿瘤外三科,宁夏 银川 750004)
乳腺癌是临床常见恶性肿瘤,目前乳腺癌发病率及死亡率逐年升高,已严重威胁人类生命安全,由于内分泌系统较为复杂导致乳腺癌早期诊断较为困难,近年来乳腺癌患者预后明显改善,但患者5年生存率仍较低,而术后乳腺癌细胞恶性增殖、迁移及侵袭是患者发生远处转移的重要原因〔1〕。因而深入探究乳腺癌发生转移的机制,积极寻找乳腺癌靶向治疗的靶点基因十分重要。微小RNA(miRNA)可通过靶向调控下游靶基因表达进而调控机体细胞各种生理病理过程,研究表明miRNA表达异常可参与多种恶性肿瘤发生及发展过程〔2〕。miR-509-3p在头颈鳞状细胞癌细胞系中表达量明显下调,上调miR-509-3p表达可通过调控表皮生长因子受体(EGFR)/磷脂酰肌醇-3-激酶(PI3K)/蛋白激酶B(AKT)/哺乳动物雷帕霉素靶向基因(MTOR)信号通路进而抑制细胞增殖〔3〕。miR-509-3p在肝癌组织中的表达低于癌旁组织,上调miR-509-3p表达可抑制肝癌细胞增殖及侵袭能力〔4〕。miR-509-3p通过下调X连锁凋亡抑制剂而抑制胶质瘤细胞增殖及侵袭〔5〕。但miR-509-3p在乳腺癌致病机制中的研究尚未见报道。长链非编码RNA CASC2可通过沉默B淋巴细胞瘤(BCL)2而增强小檗碱诱导的结肠直肠癌细胞凋亡能力〔6〕。人参皂苷-Rg5通过抑制BCL2表达进而抑制视网膜母细胞瘤增殖并诱导细胞凋亡〔7〕。starBase靶基因预测出BCL2基因可能是miR-509-3p的靶基因,由此推测miR-509-3p是否可通过靶向调控BCL2基因表达而影响乳腺癌细胞增殖、迁移、侵袭及凋亡。因此,本研究主要验证上述推测,为揭示乳腺癌发生转移机制奠定理论基础。
1 材料与方法
1.1一般资料 收集2016年2月至2017年3月于宁夏医科大学总医院就诊的乳腺癌患者65例为研究对象,患者均经手术病理证实为乳腺癌,年龄为50~70岁,平均为(65.32±7.16)岁,纳入标准:生存时间超过6个月;术前未接受化疗或放疗等治疗;临床资料完整者。排除标准:合并其他恶性肿瘤者;心肌梗死等心血管疾病病史者;依从性较差者。经医院伦理委员会批准,患者及其家属知情且签署同意书。患者均于术中切除乳腺癌组织及癌旁组织并迅速放入液氮中,术后将其转移至-80℃超低温冰箱保存备用。
1.2材料与试剂 乳腺上皮细胞MCF-10A与乳腺癌细胞MCF-7、T47D、MDA-MB-231、MDAMB-453、Hs578均购自中国科学院上海生命科学研究院细胞资源中心。DMEM培养基购自美国Hyclone公司;胎牛血清与胰蛋白酶均购自美国Gibco公司;miR-509-3p模拟物(mimics)、阴性对照miR-NC均购自Dharmacon公司(上海睿铂赛生物科技有限公司代理);Lipofectamine2000与Trizol试剂均购自美国Invitrogen公司;双荧光素酶报告基因检测试剂盒购自美国Promega公司;Transwell小室购自美国Corning公司;噻唑蓝(MTT)检测试剂盒购自上海碧云天生物技术有限公司;Mgteigel基质胶购自美国BD公司;细胞凋亡检测试剂盒购自武汉艾美捷科技有限公司;Trizol、反转录及qRT-PCR试剂盒均购自日本TaKaRa公司。
1.3方法
1.3.1细胞转染及分组 取对数生长期乳腺癌MDA-MB-231细胞,放入不含血清及青链霉素的DMEM培养基内培养,待细胞融合度达到50%时进行转染,严格按照Lipofectamine2000转染试剂盒说明书进行操作,将miR-509-3p mimics、miR-NC、anti-miR-509-3p、anti-miR-NC分别转染至乳腺癌MDA-MB-231细胞,分别为miR-509-3p组、miR-NC组、anti-miR-509-3p组、anti-miR-NC组,同时将pcDNA-BCL2与miR-509-3p mimics共转染至乳腺癌MDA-MB-231细胞,miR-509-3p mimics与pcDNA-BCL2共转染至乳腺癌MDA-MB-231细胞,分别为miR-509-3p+pcDNA-BCL2组、miR-509-3p+pcDNA组,转染6 h后更换培养基为含有10%胎牛血清的DMEM完全培养基,继续培养48 h。
1.3.2qRT-PCR检测细胞中miR-509-3p、BCL2 mRNA表达水平 收集乳腺癌组织及癌旁组织,液氮迅速研磨,加入1 ml Trizol试剂,按照 Trizol法提取组织RNA,(收集各组细胞采用Trizol法提取细胞总RNA),按照反转录试剂盒将RNA反转录为cDNA,根据qRT-PCR试剂盒说明书配置qRT-PCR反应体系,反应条件:95℃ 2 min循环1次,(95℃ 30 s,60℃30 s,72℃ 30 s)循环40次,采用2-ΔΔCt法计算miR-509-3p、BCL2 mRNA相对表达量。
1.3.3MTT检测细胞增殖 收集对数生长期乳腺癌MDA-MB-231细胞,调整细胞浓度5×104个/ml接种于96孔板(1×104个细胞/孔),放入37℃恒温培养箱继续培养,每孔分别加入20 μl MTT溶液(质量分数5 mg/ml),继续培养4 h,弃上清,加入二甲基亚砜(DMSO)150 μl/孔,低速振荡10 min,分别于溶解后的24 h、48 h、72 h时置于酶标仪检测490 nm处各孔吸光度值(OD)。实验均设置3次重复。
1.3.4流式细胞术检测细胞凋亡 用预冷磷酸盐缓冲液(PBS)洗涤各组对数生长期乳腺癌MDA-MB-231细胞,低温条件下1 000 r/min转速离心5 min(离心半径6 cm),PBS洗涤,再次离心,弃上清,分别加入5 μl膜联蛋白(Annexin)V-FITC与碘化丙啶(PI),充分混匀后室温避孵育20 min,应用流式细胞仪检测细胞凋亡率。
1.3.5Transwell实验检测细胞迁移及侵袭 取100 μl基质胶稀释液(无血清DMEM培养基:基质胶=1∶8)铺于Transwell小室上室,500 μl含10%胎牛血清的DMEM培养基加入Transwell小室的下室,继续培养24 h,擦去未侵入细胞表面的细胞,多聚甲醛固定20 min,结晶紫(0.1%)染色10 min,干燥,显微镜下拍照并计算侵袭细胞数。细胞迁移实验:取100 μl不含血清的DMEM培养基放入Transwell小室的上室,另取500 μl含10%胎牛血清DMEM培养基加入Transwell小室的下室,其余步骤同细胞侵袭实验,显微镜下观察迁移细胞数。实验设置3次重复,每组实验均设置3个复孔。
1.3.6Western印迹检测细胞周期蛋白(Cyclin)D1、基质金属蛋白酶(MMP)-2、MMP-9、p21、BCL2蛋白表达 收集各组乳腺癌MDA-MB-231细胞,加入蛋白裂解液〔1 ml RIPA、1% 苯甲基磺酰氟(PMSF)、0.1% 抑肽酶〕,冰上裂解30 min提取细胞总蛋白,二喹啉甲酸(BCA)法定量蛋白,取20 μg蛋白进行十二烷基硫酸钠(SDS)-聚丙烯酰胺凝胶电泳(PAGE)反应,蛋白凝胶转移至聚偏氟乙烯(PVDF)膜,用5%脱脂奶粉封闭1 h,孵育一抗(稀释比1∶1 000)4℃条件下孵育24 h,TBST洗膜3次,每次5 min,室温条件下孵育二抗(稀释比1∶5 000),孵育时间1 h,TBST洗膜3次,每次5 min,滴加电化学发光(ECL)剂,用Image J软件分析条带灰度值。
1.3.7双荧光素酶报告基因实验验证miR-509-3p、BCL2 的靶向关系 starBase预测miR-509-3p靶基因可能为BCL2,构建含有miR-509-3p与BCL2的结合位点的野生型载体BCL2-WT,构建含有miR-509-3p与BCL2突变结合位点的突变型载体BCL2-MUT,同时将乳腺癌MDA-MB-231细胞接种于24孔板,细胞汇合度达到80%转染,共转染分组依据:BCL2-WT与miR-509-3p mimics共转染;BCL2-WT与miR-NC共转染;BCL2-MUT与miR-509-3p mimics共转染;BCL2-MUT与miR-NC共转染,放入37℃恒温培养箱继续培养24 h,分别加入细胞裂解液(60 μl/孔),低速振荡20 min,根据双荧光素酶报告基因检测试剂盒检测各组细胞荧光素酶活性,严格按照试剂盒说明书进行操作。
1.4统计学处理 采用SPSS21.0软件进行独立样本t检验、单因素方差分析、LSD-t检验。
2 结 果
2.1乳腺癌组织及细胞系中miR-509-3p与BCL2的表达 与癌旁组织相比,乳腺癌组织中miR-509-3p表达水平显著降低,而BCL2 mRNA表达水平显著升高(P<0.05);相对于MCF-10A细胞,乳腺癌细胞MCF-7、T47D、MDA-MB-231、MDAMB-453、Hs578中miR-509-3p表达水平显著降低(P<0.05),而BCL2 mRNA表达水平显著升高(P<0.05),其中miR-509-3p在乳腺癌MDA-MB-231细胞中的表达水平降低幅度最显著,因此后续实验中选取乳腺癌MDA-MB-231细胞为研究材料,见表1、表2。

表1 乳腺癌组织中miR-509-3p与BCL2的表达
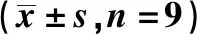
表2 乳腺癌细胞系中miR-509-3p与BCL2的表达
与MCF-10A相比:1)P<0.05
2.2上调miR-509-3p表达对乳腺癌MDA-MB-231细胞增殖及凋亡率的影响 与miR-NC组相比,miR-509-3p组乳腺癌MDA-MB-231细胞增殖活性显著降低(P<0.05),而细胞凋亡率显著增加(P<0.05),CyclinD1蛋白水平明显降低(P<0.05),而p21蛋白水平显著升高(P<0.05),见图1、图2、表3。

图1 流式细胞术检测各组细胞凋亡

1~3:空白对照组、miR-NC组、miR-509-3p组;图3同图2 Western印迹法检测各组细胞增殖相关蛋白表达
2.3上调miR-509-3p表达对乳腺癌MDA-MB-231细胞迁移及侵袭的影响 与miR-NC组相比,miR-509-3p组乳腺癌MDA-MB-231细胞迁移及侵袭细胞数均显著减少(P<0.05),MMP-2、MMP-9蛋白水平显著降低(P<0.05),见表3、图3。
表3 各组细胞增殖率、凋亡率及迁移、侵袭能力比较
与miR-NC组相比:1)P<0.05

图3 上调miR-509-3p表达对乳腺癌MDA-MB-231细胞迁移及侵袭相关蛋白表达的影响
2.4miR-509-3p靶向调控BCL2 starBase网站预测miR-509-3p的靶基因,结果显示BCL2的3′UTR中含有与miR-509-3p互补的核苷酸序列,见图4。双荧光素酶报告基因实验结果显示,与共转染miR-NC、BCL2-WT组相比,miR-509-3p mimics与BCL2-WT共转染组细胞荧光素酶活性显著降低(P<0.05);与共转染miR-NC、BCL2-MUT组相比,miR-509-3p mimics与BCL2-MUT共转染组细胞荧光素酶活性变化无显著性差异(P>0.05),见表4。进一步探究二者间调控关系,结果显示,与miR-NC组(0.88±0.12)相比,miR-509-3p组(0.45±0.10)细胞中BCL2蛋白水平显著降低(P<0.05),相对于anti-miR-NC组(0.71±0.11),anti-miR-509-3p组(1.16±0.09)细胞中BCL2蛋白水平显著升高(P<0.05),见图5。

图4 BCL2的3′UTR中含有与miR-509-3p互补的核苷酸序列


组别BCL2-WTBCL2-MUTmiR-NC组1.03±0.111.08±0.07miR-509-3p组0.43±0.051.07±0.11t/P值14.897/0.0000.230/0.821
2.5BCL2过表达逆转上调miR-509-3p表达对乳腺癌MDA-MB-231细胞增殖、凋亡、迁移及侵袭的作用 与miR-509-3p+pcDNA组相比,miR-509-3p+pcDNA-BCL2组乳腺癌MDA-MB-231细胞增殖活性显著升高(P<0.05),迁移及侵袭细胞数显著增加(P<0.05),MMP-2、CyclinD1蛋白水平显著升高(P<0.05),而细胞凋亡率显著降低(P<0.05),见图6、表5。

1~5:miR-NC组、miR-509-3p组、miR-509-3P+pcDNA组、miR-509-3P+pcDNA-BCL2组图6 Western印迹法检测各组乳腺癌MDA-MB-231细胞中相关蛋白表达
表5 各组细胞增殖、凋亡、迁移及侵袭能力比较
与miR-NC组相比:1)P<0.05;与miR-509-3p+pcDNA组相比:2)P<0.05
3 讨 论
miRNA异常表达可参与多种疾病发生及发展过程,研究表明miRNA在多种恶性肿瘤中均存在异常表达,并可通过影响肿瘤细胞增殖、迁移及侵袭等生物学过程进而影响肿瘤发生及发展过程〔8〕。研究报道指出miR-940、miR-124等在乳腺癌细胞中表达降低,上调其表达可通过抑制靶基因表达进而调控乳腺癌细胞增殖、迁移及侵袭过程〔9,10〕。但仍有部分miRNA在乳腺癌发生及发展过程中的作用机制尚未完全阐明,因此本研究主要寻找新miRNA分子与乳腺癌细胞增殖、迁移及侵袭等生物学行为的相关性,为寻找乳腺癌靶向治疗提供潜在靶点基因。
miR-509-3p通过抑制X连锁凋亡抑制蛋白(XIAP)表达进而抑制胃癌细胞增殖及迁移〔11〕。miR-509-3p可减弱卵巢癌细胞迁移及增殖能力〔12〕。miR-509-3p通过靶向丝裂原活化蛋白激酶8表达进而抑制肾细胞癌细胞增殖及迁移〔13〕。既往研究表明 miR-509-3p靶向细胞周期依赖蛋白激酶(CDK)2、Rac1等多种生长调节基因,可能有助于有效的抗癌治疗〔14〕。本研究结果表明乳腺癌组织及细胞中miR-509-3p的表达下调,与上述文献报道结果相似,进一步研究显示上调miR-509-3p表达可明显降低乳腺癌细胞活性,并可减少迁移及侵袭细胞数,而促进细胞凋亡,为验证其作用可能机制,检测细胞增殖、迁移及侵袭相关蛋白表达,结果显示CyclinD1、MMP-2、MMP-9表达水平显著降低,而p21表达水平显著升高,已有研究报道指出CyclinD1可通过与CDK4/6结合形成依赖于细胞周期蛋白的蛋白激酶(Cyclin-CDK)复合物进而正向调控细胞周期进程,而p21可通过抑制Cyclin-CDK复合物形成进而抑制肿瘤细胞周期进展,抑制细胞增殖,MMP-2、MMP-9作为基质金属蛋白酶家族成员,其可通过降解细胞外基质沉积进而促进细胞发生转移〔15,16〕。提示上调miR-509-3p表达可通过调控乳腺癌细胞增殖、迁移、侵袭及凋亡过程进而参与乳腺癌发生及发展过程。
长链非编码(Lnc)RNA H19可通过靶向调控BCL2基因表达进而影响宫颈癌细胞生物学行为〔17〕。miR-204过表达可能通过抑制BCL2基因表达进而抑制卵巢癌细胞增殖并诱导细胞凋亡〔18〕。BCL2基因表达异常涉及细胞增殖、肿瘤形成等多种过程,并可直接影响肿瘤细胞迁移及侵袭等生物学行为〔19,20〕。本研究结果显示BCL2在乳腺癌组织及其细胞中的表达水平增高,提示BCL2基因表达水平升高可能促进乳腺癌发生。靶基因预测显示BCL2基因可能是miR-509-3p的靶基因,通过双荧光素酶报告基因实验证明miR-509-3p可靶向结合BCL2基因并可负向调控其表达,进一步研究显示上调BCL2基因表达可逆转miR-509-3p过表达对乳腺癌细胞增殖、迁移及侵袭的抑制作用及其对细胞凋亡的促进作用。提示上调miR-509-3p表达可通过抑制BCL2基因表达进而减弱乳腺癌细胞增殖、迁移及侵袭能力,诱导细胞凋亡。
综上所述,miR-509-3p在乳腺癌组织及细胞中低表达,而BCL2基因高表达,上调miR-509-3p表达可通过下调靶基因BCL2表达进而抑制细胞增殖、迁移及侵袭并促进其凋亡,本研究结果证实miR-509-3p在乳腺癌发生转移过程中发挥重要作用,可丰富乳腺癌细胞转移的分子机制,可为临床乳腺癌基因治疗提供潜在靶点。
