牧草辐射诱变育种的研究进展
2019-12-09孙小富王雷挺赵丽丽黄莉娟简忠岭王家豪
孙小富, 王雷挺, 赵丽丽, 黄莉娟, 简忠岭, 王家豪
(贵州大学 动物科学学院 草业科学系, 贵州 贵阳 550025)
辐射诱变育种是人为使用γ射线、Χ射线、β射线、紫外线或中子、激光和离子束等物理因素诱导植物基因突变,促进基因重组,在短时间内得到可直接利用或有价值的突变体,从而获得新的遗传资源[1-3]。辐射诱变育种在世界上起源较早,研究也在进一步深入。我国辐射育种起步较晚,20世纪50年代末开始迅速发展,主要探讨γ射线和中子辐照处理植物,获取有经济价值的突变体。但对辐射诱变的机理、突变材料的生理生化机制以及突变体中染色体的变异行为等还有待深入探讨。近年来,随着现代畜牧业的推进和国家重点草原建设工程的实施,各地对不同用途牧草品种的需求急剧增加,为牧草育种带来了较大的发展机遇。当前,牧草育种的手段根据变异来源可分为杂交育种、诱变育种、单倍体育种和多倍体育种。根据生物工程原理可分为基因工程育种、细胞工程育种和植物激素育种。其中,诱变育种中的辐射诱变育种与其他育种手段相比,具有育种速度快、变异多和育成品种较稳定等特点[4]。为牧草辐射诱变育种提供理论基础与借鉴参考,从诱变源的种类、诱变育种特点与机制、诱变材料的选择及其诱变效应和突变体的筛选与鉴定等方面对牧草辐射诱变育种的研究进展进行了概述。
1 牧草辐射诱变源的种类
辐射诱变育种中诱变源种类主要有电离辐射和非电离辐射两大类。其中,电离辐射源主要有中子、γ射线、X射线和β射线,非电离辐射主要有紫外线、离子束和激光。中子为不带电的粒子,半径约为8×10-15m,与质子大小类似;应用材料有玉米[5]、紫花苜蓿[6]和红三叶[7]等。γ射线是原子核内发出的高能电磁辐射,其波长短(<0.1 nm)、穿透力强和射程远,应用材料有扁穗牛鞭草[8]、沙打旺[9]、狗牙根[10-11]、高羊茅[12]、紫花苜蓿[13-14]、二穗短柄草[15]、垂穗鹅观草[16]、白刺花[17]和红豆草[18]等。X射线波长0.01~0.1 nm,是一种高能电磁辐射,分硬X射线和软X射线2种。硬X射线能量较大,波长较短,穿透能力较强;软X射线波长较长,能量较低,但能产生较高的线性能量转移;应用材料有玉米[19]和大麦[20]等。β射线是由放射性同位素衰变时所放出的带负电荷的粒子流,质量小,在空气中射程远,穿透力弱,辐射源为同位素32P和35S;应用材料有小麦[21]。紫外线波长250~290 nm,是一种穿透力小的非电离射线,为低能电磁辐射;应用材料有中华羊茅、冷地早熟禾和垂穗披碱草[22]、狭叶披碱草和红毛羊胡子草[23]、多花黑麦草[24]和龙须草[25]等。离子束是由元素的离子经高能加速器加速后获得,对植株生理损伤小,突变频率高,具有一定的方向性和重复性;应用材料有甜菊[26]和拟南芥[27]等。激光亮度高、单色性、方向性和相干性均好,属低能电磁辐射;应用材料有高羊茅[28]。
2 牧草辐射诱变育种特点
2.1 提高突变率,扩大变异范围
在自然条件下,由于外界环境的变化和遗传结构的不稳定性,植物会产生自然突变。然而,这种突变的频率非常低,而人工诱导突变因子引起的突变频率比自发突变要高数百倍,甚至数千倍,变异幅度大,类型多样,为新品种选育创制新的种质资源[29]。谢向誉等[30]利用不同剂量60Co-γ射线辐照新选048和华南205 2个木薯品种,采用SCoT分子标记分析检测得出,辐射剂量为50 Gy和100 Gy时,华南205的表型变异率较对照组(0 Gy)分别提高1.2%和3.5%,新选048的表型变异率较对照组(0 Gy)分别提高1.5%和5.4%。张彦芹等[31]利用不同辐射剂量60Co-γ射线对爱瑞3号高羊茅干种子和分化苗进行辐照处理表明,在诱变剂量为100~150 Gy时,干种子变异率为1.52%~8.33%,在诱变剂量为20~25 Gy时,分化苗变异率为1.52%~1.85%。
2.2 打破性状连锁,改良个别单一性状
在牧草品种中,有益性状和不良性状常会出现连锁现象。辐射诱变育种可使个别基因位点引起单个性状变异,将多个性状分开,改良品种中的不良性状。在进行早熟、抗病、矮秆和品质育种时,辐射诱变育种的效果较好,同时能够保持该品种原有的其他优良特性[29]。刘勇等[32]报道,利用不同剂量60Co-γ射线辐照处理黑麦草,可显著提高其产量。鱼红斌等[33]利用辐射诱变育种技术促使沙打旺生育期缩短,培育出早熟沙打旺品种。
2.3 诱发变异较稳定,缩短育种年限
辐射可以提高植物遗传变化的机率,且产生的变异多数是1个主效基因的改变,经3~4代选育基本达到稳定,从而培育出新的种质资源[29]。特别是利用辐射诱变育种手段与组织培养技术相结合,可有效提高变异的保存率,加快变异的纯合,缩短育种周期。同时,对植物细胞进行辐射诱变时,发生单细胞突变的再生苗一般为纯合突变体,因而可提高育种效率,缩短育种周期。马建中等[34]对沙打旺草籽进行辐照处理表明,辐照处理可提高其突变体的适应性和选择的准确性,缩短育种周期。
2.4 改变植物育性,促进远缘杂交
利用辐照处理远缘杂交材料的花粉,可促进受精结实,克服杂种不育。此外,辐照处理能使正常的植株产生雄性不育,为雄性不育利用提供原材料[29]。20世纪中叶,人类在开展黑麦草和羊茅属牧草进行杂交时,利用γ射线辐照处理,能够极大的减轻杂交不亲和性,最终选育出既有黑麦草性状又具有羊茅属牧草耐高温干旱性状的杂交牧草新品种[35]。在对自交不亲和的白三叶进行杂交育种时,利用辐射诱变育种手段能够极大地降低自交不亲和性[30]。
3 辐射诱变育种的作用机制
3.1 细胞水平
辐射诱变育种的机理研究主要围绕染色体畸变和突变关系展开。染色体畸变是植物辐射损伤的典型特征,包括染色体数目和结构两种变异类型,染色体数目变异包括倍性变异和非倍性变异,染色体结构变异包括染色体的修复和重新连接后的易位、倒位和缺失等[36-37]。李健等[38]利用不同剂量60Co-γ射线辐照处理野牛草种子表明,γ射线辐照能诱发野牛草根尖细胞的染色体发生畸变,抑制野牛草根尖细胞的有丝分裂,且随辐射剂量的增加,根尖细胞有丝分裂指数呈下降趋势。
3.2 分子水平
诱变机理研究主要围绕DNA的损伤、修复与突变形成的关系进行探讨。在利用非电离辐射中的紫外线进行辐射诱变时,DNA或RNA上相邻的嘧啶以共价键相互结合形成环丁烷嘧啶二聚体结构,是紫外线生物学作用的主要原因。植物的突变与转录因子结合位点密切相关,且突变的频率在整个基因组中高度可变[39-41]。转录因子结合位点可以改变DNA中紫外线损伤的速率,转录因子结合位点极易受到紫外线的损伤,DNA中碱基序列发生突变(替换、插入等)有利于形成环丁烷嘧啶二聚体结构,且随着紫外线损伤的频率增加可能导致突变频率的提高[39,42-43]。然而,大多数转录因子通过限制形成环丁烷嘧啶二聚体结构来抑制紫外线对结合位点的损伤[44]。紫外线照射时,DNA分子所形成的TT二聚体通常不会导致突变,大多数紫外线诱导的突变是TC二聚体和CC二聚体,且C的突变频率大于T[45]。利用电离辐射中的γ射线辐照处理植物种子其发芽率呈升高或者降低变化趋势。一般情况下,低剂量的γ射线会促进种子的萌发,高剂量的γ射线抑制种子萌发和幼苗的生长,且低剂量辐照时,有利于促进种子中RNA和蛋白质的合成[46-47]。种子在高剂量γ射线下不能发芽的原因[48]:组织学和细胞学的改变,种子层和细胞膜的破坏与辐射剂量成正比,种子发芽过程中分生组织区域或有丝分裂受到破坏。辐射常会引起DNA置换突变,即碱基的替换,但由于遗传密码的简并性,同义替换后的mRNA在翻译为氨基酸时,氨基酸序列并未改变,即蛋白质的性状并未发生改变。
3.3 形态及生理水平
利用γ射线辐照植物体时,其外部形态特征和体内的各种生理生化特征发生改变[46,48]。γ射线的生物学效应基于与细胞中的原子或分子(特别是水)的相互作用产生自由基,这些自由基可破坏或改变植物细胞的重要组分,且辐射剂量不同,对植物的形态学、解剖学、生物化学和生理学产生影响也不同[46,49]。这些影响包括植物细胞结构和代谢的变化,如类囊体膜的扩张、光合作用的改变、抗氧化系统的调节和酚类化合物的积累等[50]。低剂量辐射将通过改变植物细胞中的激素信号传导网络或通过增加细胞的抗氧化能力来诱导生长刺激,增加活化酶系统来增强叶绿素的合成,高剂量辐射诱导的生长抑制归功于体细胞分裂期间细胞周期停滞在G2或M期,对整个基因组造成不同程度的损害,抑制叶绿素合成[51-53]。此外,辐射处理会引起植物体内超氧化物歧化酶(SOD)[54]、过氧化物酶(APX)[54]、过氧化氢酶(CAT)[54]、谷胱甘肽还原酶(GR)[55]和丙二醛(MDA)含量[56]等的变化。
4 辐射诱变育种的材料选择及其诱变效应
4.1 辐射处理方法
辐射处理方法可分为内照射和外照射2种。外照射指被照射的种子或植株所受的辐射来自外部某一辐射源。内照射是将辐射源引入被照射种子或植株某器官内,通常采用放射性32P和35S溶液浸种,或将32P和35S溶液注射到植株营养器官(根蘖、茎、枝条和芽等)中,或是施于土壤让植株吸收[35]。外照射处理方法具有简便、安全和效果佳等特点,在育种中应用较广泛。内照射需要一定的设备和防护条件,且易造成污染,在育种中应用较少[57]。
4.2 材料选择及其诱变效应
目前,牧草辐射诱变处理材料有活体植株、植株营养体(根蘖、茎和幼芽等)、种子和组织培养物等(表1)。活体植株辐照是将生长至某一时期植株的整体或局部置于有辐射源的辐射室内进行辐照处理。由于活体植株正处于生长旺盛时期,对辐射较为敏感,且不同发育阶段其敏感性存在较大差异。因此只有采用适量的辐射剂量才能获取较为理想的突变效果。植株营养体是牧草辐射诱变育种中常选用的诱变材料之一,在牧草辐射诱变育种中,主要应用于种子成熟困难且靠无性繁殖的牧草。种子(包括干种子、湿种子和已萌动种子)是辐射诱变中常选用的诱变材料,可用于不同的辐射诱变源[59]。其中,干种子作为诱变中常用的材料,具有代谢水平低、操作简单、携带方便、可长途运输和便于大量处理等特点,能够在干燥、高温、极低温和真空等条件下进行处理,能够忍受较高剂量的射线辐照。湿种子和已萌动的种子与干种子相比,具有对辐射诱变源更敏感,但难以保存、携带和贮藏等特点。辐射诱变处理可诱导植物体细胞无性系变异,植物体细胞无性系变异是指在植物组织培养再生植株过程中出现的变异,是植物组织培养过程中普遍存在的现象[78]。应用组织培养技术进行辐射诱变育种,诱变产生的突变可在细胞水平上直接呈现,可提高突变发生概率、增加选择机会、改变分离纯化速度和缩短育种周期,这种离体诱变技术在国内外逐渐发展起来[79]。
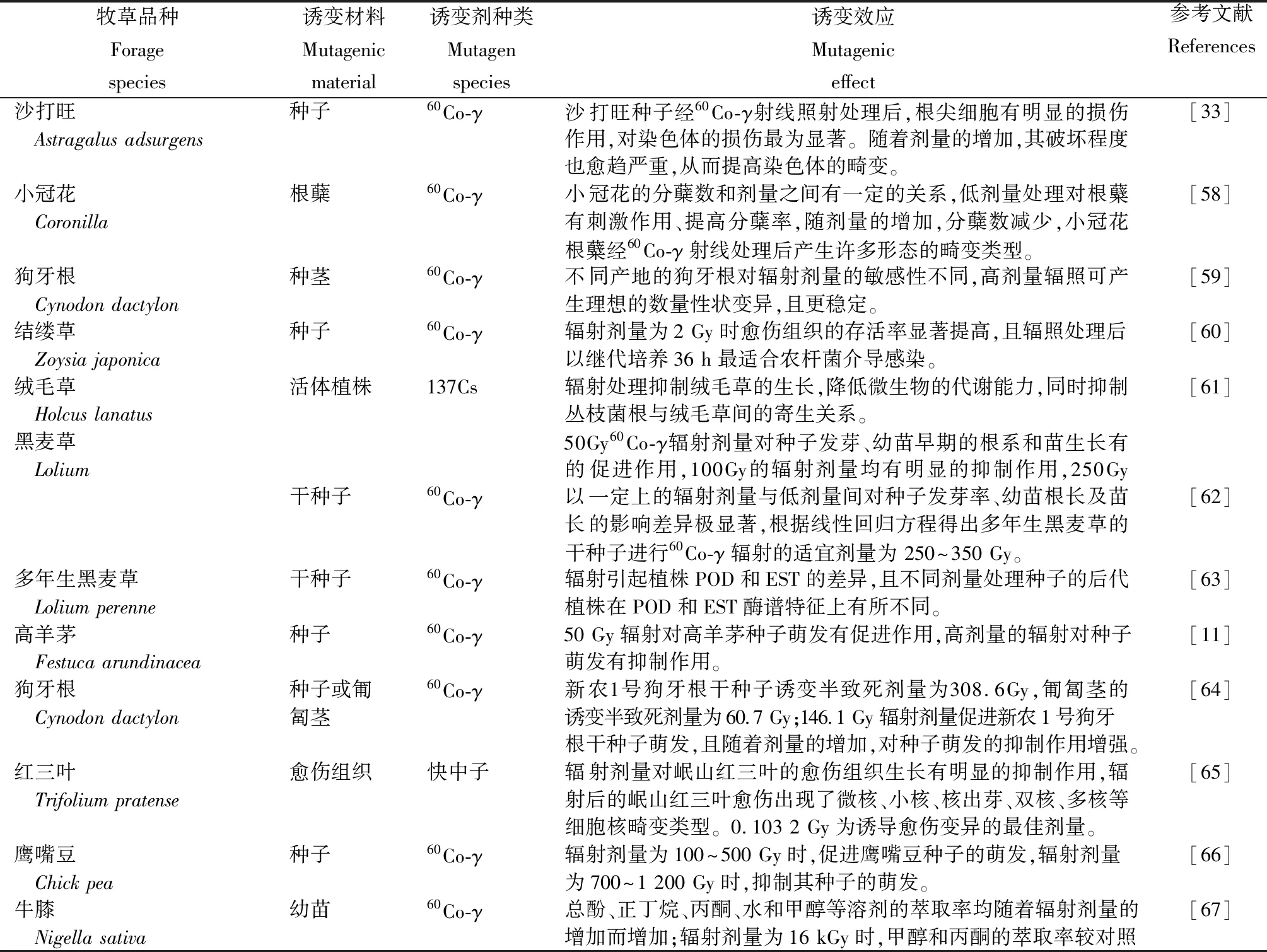
表1 牧草辐射诱变材料的选择及其诱变效应
续表1

牧草品种Forage species诱变材料Mutagenic material诱变剂种类Mutagen species诱变效应Mutagenic effect参考文献References组提高5.40%和10.60%。狗牙根种子60Co-γ狗牙根各辐照变异植株与对照之间在节间长均有显著差异,形态[12] Cynodon dactylon变异植株与其对照之间的遗传相似系数为0.585~0.776,遗传差异性为较明显。野牛草干种子60Co-γγ射线辐照能抑制野牛草根尖细胞的有丝分裂。随γ射线辐射[68] Buchloe dactyloides剂量的增加, 根尖细胞有丝分裂指数呈下降趋势;γ射线辐照能诱发根尖细胞的染色体畸变和核畸变, 产生多种畸变类型。日本结缕草干种子60Co-γ低剂量辐射处理促进日本结缕草干种子的萌发,提高其发芽率,[69] Zoysia japonica随着辐射剂量的增加,发芽率逐渐降低,临界辐射剂量为450 Gy,半致死剂量为480 Gy。红三叶干种子快中子低辐射剂量(11.01~29.31 Gy)有利于红三叶根长伸长,且对幼苗[70] Trifolium pratense出苗率和株高有一定促进作用。高剂量辐射(>29.31 Gy)可促进红三叶种子萌发,茎长生长,但对出苗率和株高均有一定抑制作用。29.31 Gy 辐射剂量最适宜岷山红三叶种子的萌发及其幼苗生长。幼苗生长。香附子种子60Co-γ在较高剂量的射线下,香附子种子萌发天数和萌发完成天数显[71] Lepidum sativum 著推迟,幼苗生长状况随着辐射剂量的增加呈下降趋势;低剂量的射线能促进其种子的萌发。中华羊茅、冷地早熟禾、活体植株UV-B随着辐射剂量的增强,牧草群体结构发生改变,群体叶面积指数[22]垂穗披碱草(LAI)下降。牧草群体对UV-B辐射增加的反射增大,吸收减少, Festuca sinensis Keng、尤其是上层叶片对UV-B辐射的吸收减少,减轻UV-B辐射的 Poa crymophila、伤害,且中下层叶片也由于上层叶片的遮挡,处于较低UV-B辐射 lymus nutans水平。沟叶结缕草茎段愈伤60Co-γ较低辐射剂量(5 Gy)对愈伤组织的生长、再生无影响,较高辐射[72] Zoysia matrella组织剂量(40 Gy)使愈伤组织的胚性保持能力和再生率下降,适当的辐射剂量(10 Gy,20 Gy)对愈伤组织的再生速度有显著的促进作用。垂穗鹅观草干种子60Co-γ辐射剂量小于或等于50 Gy时,有利于种子的萌发,50 Gy种子[16] Roegneria nutans萌发能力下降。辐射半致死剂量为148 Gy,适宜剂量为 0~148 Gy。结缕草茎段愈伤60Co-γ15~20 Gy辐射剂量处理有利于促进结缕草愈伤组织向胚性愈[73] Zoysia japonica组织伤组织转化,获取较高的出愈率和胚性愈伤组织形成率。直立型扁蓿豆干种子或60Co-γ干种子辐射后,辐射剂量为1 400 Gy时,M0代植株株型为斜生[39] Medicago ruthenica吸胀种子型,辐射剂量为1 000 Gy时,表现出荚果大、叶片小、多叶、叶绿素含量较高和生长快;在辐射剂量为800Gy时,表现出多叶和叶面积大等优良特性;吸胀种子辐射后,M0代植株在辐射剂量为1 000Gy时,表现为直立型,且花小、荚果大、种子宽、多叶、叶绿素含量较高和生长快,在辐射剂量为1200Gy时,表现出荚果宽、种子大和多叶等优良特性。扁穗牛鞭草种茎60Co-γ雅安牛鞭草茎适宜的诱变剂量为80~100 Gy,H055扁穗牛鞭[8] Hemarthria cornpressa草茎适宜的诱变剂量为70~90 Gy。紫花苜蓿干种子60Co-γ高剂量辐照对紫花苜蓿种子的诱变致畸效果显著增加。[74] Medicago sativa白刺花干种子60Co-γ随着辐射剂量的增加,白刺花的各项生长指标(叶绿素ChI、可[17] Sophora davidii溶性蛋白、可溶性糖、MDA、POD)呈现升高后降低的趋势,不同辐射剂量均可促进白刺花幼苗的光合作用。海滨雀稗种茎60Co-γ所有突变材料植株的叶宽、叶长、株高、匍匐茎节间长度和直径[75] Seashore paspalum以及密度等指标均不同程度地优于对照,突变材料SI-50-1、PL-40-2和SLM-45-1叶片短细、节间缩短、株高矮化和成坪密度高,辐射诱变效果明显。白三叶幼苗60Co-γ通过对120份材料进行诱变处理,共获得97个突变体,其中4份[76] Trifolium repens具有极强的抗旱性。象草种茎60Co-γ辐射处理后的植株由矮化的趋势,辐射较易引起象草的分蘖数、[77] Pennisetum purpureum茎节间和叶长的变异,象草种茎的适宜辐射剂量为30 Gy。
5 辐射诱变育种中突变体的选择与鉴定
经诱变处理的种子长成的植株为辐射当代(M0),利用辐射当代种子长成的植株为M1代,由M1代种子繁殖长成的植株为M2代。M1代是由诱变直接处理当代细胞衍生而来,变异具有局部性,多为复杂的突变嵌合体,一般不作选择,进行保苗即可,若辐射诱变材料为杂种、异花授粉植物或单倍体的群体,M1可能出现分离现象,应进行选择。而M2代为性状分离世代,也需要进行选择[29]。
5.1 形态学鉴定
突变体的鉴定和选择是辐射诱变育种的又一关键技术。传统农艺性状的观察和判断,即以形态学标记作为选择标准。刘天增等[75]对4个海滨雀稗品种的诱变突变体材料进行形态鉴定,筛选出9个突变体材料。贾彩凤[80]用60Co-γ射线对金荞麦的根茎进行辐射处理,根据其后代的农艺性状筛选出具有药用价值的突变体材料。辐射引起的突变分为遗传突变和非遗传突变,其中可以遗传的优良变异是育种工作需要的变异。由于许多辐射引起的形态变异(株高、叶形等)都是由生理损伤引起的,是植物可以修复的损伤,这种变异不具有遗传性,所以从形态表现直接决定突变体有其局限性。此外,表型检测受辐射后的效应、环境与基因型相互作用的影响,因此选择的结果往往也不可靠[73]。
5.2 同工酶鉴定
同工酶在辐射诱变育种中作为检测辐射生物学效应和突变体鉴定的生化指标,以过氧化物酶和酯酶同工酶的应用居多[80]。过氧化物酶作为呼吸代谢的最终氧化酶,在植物的生长发育、新陈代谢和机体适应等方面发挥着重要作用[80-81]。李永祥等[82]对辐照后的野牛草过氧化物酶和多酚氧化酶进行酶谱分析得出,幼叶条带数比幼穂多,幼穂的过氧化物酶酶谱和幼叶的多酚氧化酶酶谱分析可鉴定野牛草性别。尹淑霞[63]利用60Co-γ射线对多年生黑麦草爱神特(Accent)干种子进行辐照处理,对辐照后其植株过氧化物同工酶酶谱分析表明,辐射能够引起植株过氧化物同工酶的差异。酯酶同工酶作为一种单体酶,遗传较稳定,对突变体进行筛选时具有分析快速、简便和准确等特点。尹淑霞等[83]利用60Co-γ射线辐照处理多年生黑麦草超级德比(Derby Supreme)干种子,对植株过氧化酶(POD)和酯酶(EST)同工酶进行酶谱分析表明,辐射能够引起植株POD和EST的差异,且不同辐射剂量显著影响植株POD和EST酶谱特征。利用同工酶筛选鉴定突变体是基于蛋白质中氨基酸的变化,氨基酸的种类和含量差异导致电荷值和电性的差异,电泳时能够检测出突变体[82]。但并非所有氨基酸的变化(如氨基酸的置换)均会引起电荷的改变,利用电泳原理无法检测出该类突变体。因此,利用同工酶分析进行突变体的筛选和鉴定仍有一定的局限性[83]。
5.3 DNA分子标记鉴定
在现代分子生物学中,DNA分子标记的方法主要有扩增片段长度多态性(Amplified Fragment Length Polymorphism,AFLP)、相关序列扩增多态性(Sequence-Related Amplified Polymorphism,SRAP)、随机扩增多态性DNA (Random Amplified Polymorphic DNA,RAPD)、简单重复序列(Simple Sequence Repeat,SSR)和区间简单重复序列(Inter-simple Sequence Repeat,ISSR)等[63]。AFLP和SRAP是以PCR技术为基础,并与限制性酶切相结合的分子标记技术[63,84]。AFLP是用成对的限制性内切酶酶切基因组DNA使其产生粘性末端,粘性末端与人工接头5’连接,形成特异性引物扩增到DNA片段,利用聚丙烯酰胺凝胶电泳产生多种多态性带型的扩增片段[75,80]。冯霞[62]利用不同剂量60Co-γ 射线辐照处理多年生黑麦草种子,对M1代变异植株进行AFLP分子标记没有检测到变异位点,可能与AFLP标记的局限性和多年生黑麦草庞大的基因组有关,或是突变并非发生在核DNA。SRAP采用预先设计的特殊引物设计对进行上端外显子扩增、下端启动子与内含子区域扩增,由于个体和物种间的差异性,产生不同间隔长度的内含子、启动子,从而产生多态性[84]。严欢[85]利用不同剂量60Co-γ 射线辐照处理5份表现良好的狗牙根材料,用SRAP技术对M1代植株DNA突变效应检测结果表明,5份狗牙根材料的遗传相似系数为0.577~0.966,平均GS为0.728,遗传相似性变化范围较大,诱变材料中检测筛选出突变体。
RAPD、SSR和ISSR是以PCR技术为基础的分子标记技术[84]。RAPD是以随机脱氧核苷酸序列作为引物,通过PCR反应进行DNA片段非定点扩增,利用凝胶电泳分析DNA多态性的一种分子标记方法[30,80]。张彦芹等[31]利用60Co-γ射线辐照处理高羊茅种子和分化幼苗,对辐射当代突变体幼嫩叶片经RAPD分析筛选出DNA分子水平上发生变异的突变材料。吴立蓉等[86]利用60Co-γ 射线辐照处理广藿香外植体,利用RAPD分子标记技术对辐射当代植株分析表明,不同辐射剂量处理的外植体再生植株多态性各不相同,通过辐射处理后的外植体植株的存活率和再生能力均降低。徐国忠[87]利用60Co-γ 射线辐照处理决明属5个牧草品种种子,对M3代性状筛选鉴定表明,5个品种辐射后代在遗传上都有明显的变异,并筛选到10个在产量和营养品质等方面均优于原品种的新品系。SSR也称为微卫星序列标记(Microsatellite sequence,MS)或短串联重复标记(Short Tandem Repeat,STR),其原理是根据微卫星序列两端设计1对特殊的互补片段引物,利用PCR技术扩增卫星片段,因单元重复次数存在差异,从而形成SSR座位的多态性[88]。程晓丽[89]利用60Co-γ射线辐照处理草坪型狗牙根,利用SSR对亲本和辐射诱变后代进行分子标记表明,辐射诱变后植株花序数量的变异可能与分子标记产生的不同位点有关。ISSR是利用锚定的微卫星DNA为引物,即在SSR序列的3′端或5′端加上2~4个随机核苷酸,对SSR两侧进行反向扩增,所扩增的多条带通过电泳将其分辨出来,扩增带多为显性表现[84]。王文恩[90]采用不同辐射剂量60Co-γ 射线辐照处理3种暖季型草坪草(野牛草、狗牙根和日本结缕)干种子,利用ISSR技术对各材料间进行遗传多样性和聚类分析表明,各材料间存在较为明显的遗传差异性,筛选出的形态变异植株和对照处理间的遗传相似系数为0.540~0.780,筛选出新株系1个。
6 展望
近年来,随着国家重点草原建设工程和西部大开发战略的实施,以及生态环境建设的加强、农业产业结构的调整和现代畜牧业建设的推进,全国各地对不同用途牧草品种的需求急剧增加,牧草育种工作进入了快速发展的阶段。现阶段,我国牧草辐射育种已取得显著性进展,育种方法和手段愈趋成熟。与常规杂交育种相比,辐射诱变育种具有育种时间缩短和变异类型多样等特点,是培育牧草新品种的有效手段。通过诱变育种技术获得许多具有优良性状的突变材料,培育出爱瑞3号高羊茅(FestucaarundinaceaNo.3)[30]、闽育1号圆叶决明(ChamaecristarotundifoliaMinyu No.1)[34]、早熟沙打旺(Astragalusadsurgens)[33]、航苜1号紫花苜蓿(MedicagosativaHM-1)[91]、航苜2号紫花苜蓿(MedicagosativaHM-2)[92]、西德小冠花(Germanycrownvetch)[93]和白花三叶草(Trifoliumrepense)[94]等许多牧草新品种,使我国的牧草产业取得较大的发展。但辐射诱变育种的研究与应用仍存在以下问题,是科学工作者不断探索研究的重点。
第一,进一步加强对辐射诱变育种机理的研究。目前,我国牧草辐射诱变育种仍处于初级发展阶段,在牧草诱变机理、提高突变预见性和选择鉴定等方面的研究不够深入。辐射诱变的机理尚不完全清楚,对突变体的遗传特性、突变材料的遗传控制基因以及突变体的代谢途径研究不够透彻,加强对以下几个方向的研究,有利于帮助人类理解植物对外界环境的适应机制,明确辐射诱变的作用机理。一是“辐射后植株的形态解剖、生理生化特性、染色体的结构变异以及DNA的损伤、修复与突变形成的相关性”,二是“不同剂量辐照使植株DNA的损伤、修复与突变形成的关系”,三是“不同剂量辐照处理植物体内各种生理生化反应以及各种信号转导机制的变化”,四是“辐射诱变因素对植物生长发育规律的影响”。
第二,辐射诱变育种应与传统的育种方式相结合,培育出遗传稳定的牧草新品。单一的辐射诱变育种在培育新品种时较为困难,应与杂交育种、倍性育种和细胞工程育种等相结合,应用先进的分子技术加强对辐射突变材料的选择和鉴定,加快牧草育种的进度。同时,利用多种诱变因素进行复合诱变可能会取得较好的效果,应充分利用太空诱变的优势开展诱变育种工作。
第三,充分发挥辐射诱变植物中次生代谢物及新基因的作用。辐射诱变植物中次生代谢物可诱导产生高价值、低成本的抗肿瘤及抗癌等化合物,在生产实践中具有较大应用前景。同时,诱变产生的突变体可以用于新基因发现和功能基因组学的研究。
第四,加强诱变牧草新品种的示范推广,促进草产业的发展。辐射诱变育种作为一种新的育种方式,育成的新品种具有在短期内迅速适应不同地点、不同农业气候条件和不同生长条件等特点,但在实际生产上很难得到大面积的推广与利用。在现有资源有限的情况下,新品种在草产业及畜牧业中的作用未能得到充分发挥。建议科研机构与企业合作,加速田间试验和大面积推广,进一步使科研、示范与推广实现一体化发展,进而实现诱变育种的产业化发展。
