SJIR-2基因真核表达载体的构建及在真核细胞内表达的研究
2019-12-02王正印尹元陆群
王正印 尹元 陆群
血吸虫病是严重危害发展中国家人民健康的重要寄生虫病。全球有78个国家和地区流行血吸虫病,感染人口接近3亿,其中适龄儿童约占46%[1],为了实现消除血吸虫病的目标,研发血吸虫病疫苗成为主要的发展方向之一[2]。在血吸虫基因工程疫苗(包括核酸疫苗)研究中,迄今已筛选出多个亚单位候选抗原,可对血吸虫病的防治起到一定的作用[3~5]。为了研究更有保护力的血吸虫疫苗,本文从血吸虫胰岛素受体基因(Schistosoma ja-ponicuminsulin receptor-2,SJIR-2)方面着手,通过构建SJIR-2基因的真核表达载体作为重组疫苗,以脂质体介导的方法将重组SJIR-2基因导入293T细胞,再以Western blot的方法验证其在真核细胞内的表达,这将为进一步探索SJIR-2基因的重组疫苗在血吸虫病防治中的作用提供帮助。
材料和方法
一、材料
1.日本血吸虫、真核表达载体和pEGFP 293T细胞均来自于安徽医科大学。
2.主要试剂及仪器
Lipo2000和Trizol试剂购自美国invitrogen公司,rTaq mix购自日本TaKaRa公司,逆转录试剂盒购自美国Fermentas公司,T4连接酶、XmaI和PstI内切酶购自北京NEB公司,DNA胶回收试剂盒购自德国QIAGEN公司,质粒提取试剂盒购自美国Axygen公司,HRP标记山羊抗鼠IgG购自北京全式金公司,鼠抗pEGFP购自英国Abcam公司。
二、方法
1.RT-PCR
将日本血吸虫研碎,用Trizol提取日本血吸虫的总RNA,逆转录成cDNA,以cDNA为模板,SJIR-2基因扩增引物SJIR-2-F:GCTGCAGTATGCTAAATATATTGGCTCAACATG;SJIR-2-R:CCCCGGGATAAGCTAAATCTCCATTTGTAATTGTT进行PCR反应,反应条件是:94℃预变性5 min,接着94℃变性1 min,58℃退火1 min,72℃延伸1 min,最后72℃延伸10 min,4℃停止,共35个循环。取5μL PCR产物,进行琼脂糖凝胶电泳,用DNA胶回收试剂盒回收纯化扩增产物。
2.PCR产物和载体pEGFP的酶切及纯化
取pEGFP载体和PCR产物各10μL,用限制性内切酶PstI和XmaI进行双酶切反应,回收纯化酶切DNA片段。
3.SJIR-2基因与pEGFP的重组与鉴定
将pEGFP载体和纯化后的SJIR-2基因以1∶3的比例混合,然后加入1μL T4连接酶制成连接反应体系,将该连接反应体系放进16℃温箱孵育过夜。次日,将连接产物转化到大肠杆菌DH5α感受态内,在卡那抗性培养平板上培养12~16 h,挑取若干单克隆放入3 mL卡那霉素抗性的LB液体培养基中37℃,200 rpm振摇过夜,提取重组质粒。分别采用PCR法、双酶切法和测序法鉴定重组基因,同时进行Blast比对分析。
4.SJIR-2基因在真核细胞内的表达
以脂质体介导的方法分别将pEGFP-SJIR-2重组质粒和空pEGFP对照转染到293T细胞内,具体方法如下:①取两个1.5 mL的EP管,向两个EP管中加入50μL Opti-MEM培养基,然后向其中一个EP管中加入0.8μg质粒,向另一个EP管中加入2μL Lipofectamine 2 000,混匀后室温静置5 min。②将两个EP管中的溶液混合,室温静置20 min。③取细胞密度在90%左右的24孔板,弃掉原有的培养基,向每孔加入400μL不含血清和抗生素的DMEM培养基,将②中质粒和Lipofectamine 2000的混合液加入培养板孔中。④将24孔板放回37℃,5%CO2细胞培养箱中培养4~6 h后更换完全培养基。继续培养24 h后用荧光显微镜观察转染情况,提取转染后细胞的总蛋白,Western blot检测SJIR-2基因所编码的蛋白表达情况。
结 果
一、目的基因SJIR-2 PCR扩增结果
PCR法扩增目的基因得到1575 bp的基因片段,电泳结果与预期一致(图1)。

图1 目的基因SJIR-2的PCR扩增产物凝胶电泳分析
二、重组载体pEGFP-SJIR-2的鉴定
用PstI和XmaI双酶切重组基因可得到2个片段,即1 575 bp处的目的基因片段和约4 700 bp处的pEGFP片段(图2),将酶切得到的目的基因片段进行测序,测序结果经Blast比对与SJIR-2基因序列同源性为100%。

图2 pEGFP-SJIR-2的PCR及双酶切鉴定结果
三、SJIR-2蛋白在真核细胞内的表达
用LiPo 2000分别介导pEGFP和pEGFP-SJIR-2质粒转染293T细胞,24 h后荧光显微镜下观察到绿色荧光(图3);Western blot检测出目的蛋白的表达,与预测大小一致(图4)。
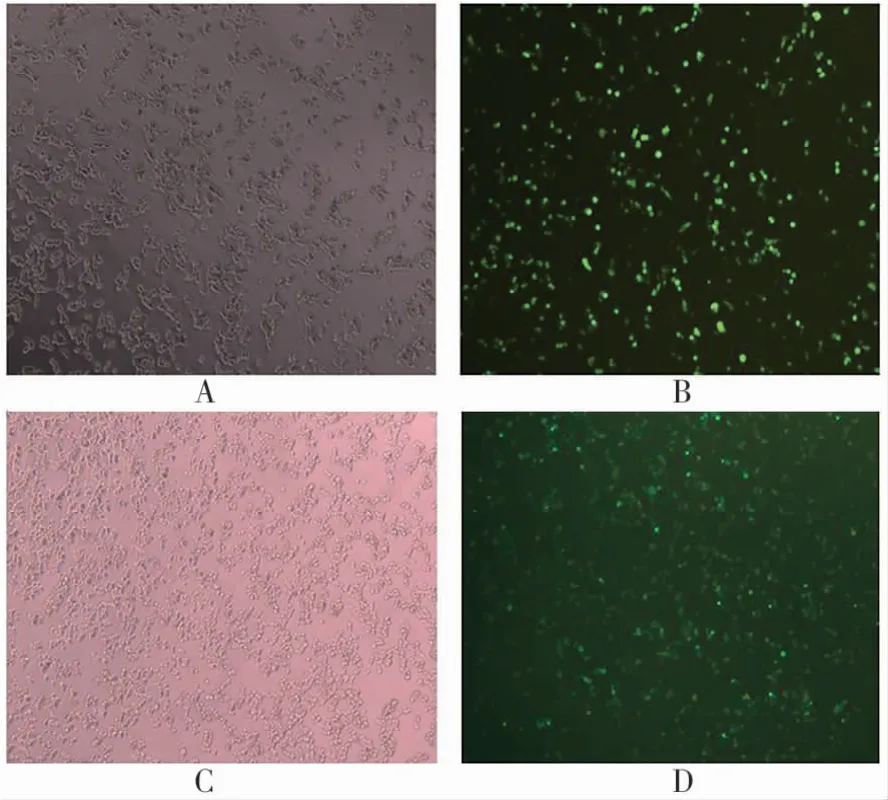
图3 荧光显微镜下结果

图4 pEGFP和pEGFP-SJIR-2质粒转换293T细胞SJIR-2 Western blot蛋白表达结果
讨 论
研制有效的血吸虫病疫苗是预防和控制血吸虫病的必要措施之一[6]。近年来,虽然在血吸虫疫苗的研究方面取得了一定的成绩[7],但是至今都没有理想的血吸虫疫苗问世。葡萄糖和糖原等碳水化合物被认为是吸虫重要的能量来源,葡萄糖和糖原等碳水化合物的代谢离不开胰岛素。研究表明[8]胰岛素介导糖代谢主要是通过PI3K途径发挥作用:胰岛素首先与靶细胞表面的胰岛素受体(IR)结合,激活IRp的酪氨酸激酶,然后导致一系列IRS的磷酸化,磷酸化的IRS激活P13K,进而引起一系列蛋白质磷酸化级联反应,最终导致胰岛素的生物效应。其中任何一个环节改变都可致胰岛素生物效应降低而产生胰岛素抵抗。因为靶细胞膜上的胰岛素受体只需10%~20%与胰岛素结合,便足以发挥最大生理效应,故胰岛素受体后信号转导障碍是胰岛素抵抗的主要原因[9]。血吸虫胰岛素受体和人类胰岛素受体在配体区域有相同的结合机制,在酪氨酸激酶区域有一样的下游信号转导加工[10]。研究表明,血吸虫除了能应答自身内在的内分泌激素,也能接受宿主激素信号调控增殖、发育和交配[11]。日本血吸虫和其哺乳动物宿主分享胰岛素受体或胰岛素样生长因子受体的基因片段,能接受宿主激素信号调控自身的发育[12,13]。另外在血吸虫童虫和成虫表面表达的膜蛋白是疫苗和药物研制合理的目标[14],而胰岛素受体就是膜蛋白,可以作为疫苗的抗原分子。本次研究成功构建了pEGFP-SJIR-2重组质粒,并且验证了其在真核细胞内的表达,从而可将该重组质粒通过一定的方法导入动物体内,使该基因编码的蛋白在宿主体内表达,诱导宿主产生特异性的抗体从而达到预防和治疗血吸虫病的目的。这些都为我们进一步探索SJIR-2基因的重组疫苗在血吸虫病防治中的作用打下坚实基础。
