棕果蝠胃及肠道的解剖与组织结构的初步观察
2019-11-29刘为民林俊青罗来福叶亚琼赵海全吕宗吉
刘为民,林俊青,罗来福,叶亚琼,赵海全,张 晖,陈 芳,吕宗吉
(佛山科学技术学院生命科学与工程学院,广东佛山528231)
蝙蝠是哺乳动物中真正能够飞翔的类群,其数量和种类仅次于啮齿类动物[1]。除两极以外,世界上几乎所有地方均有分布,但以热带地区最为丰富。果蝠是其中体型较大的一类,属于狐蝠科,以龙眼、香蕉、荔枝、各种花蕊以及甘蔗为食[2-4],生活于印度、泰国、缅甸等东南亚国家以及我国的广东、厦门、海南、广西等热带地区。近20余年,由于蝙蝠作为多种烈性传染病的病原贮存宿主而受到研究者甚至全社会的广泛关注[5-7]。但对于蝙蝠的基础研究资料还相当稀少,其本身的结构特点与病原的关系,甚至其免疫机能与结构的关系均是当前亟待深入研究的的领域。
本研究采用大体解剖与组织学相结合的方法,初步观察了生活在广东省云浮市境内的棕果蝠的胃肠道结构,以便了解这类适应飞翔的哺乳动物其结构发生了哪些适应性变化,为后续的相关研究提供基础性支撑成果。
1 材料与方法
棕果蝠5只,体重100~120 g,均为雌性,采集自云浮山区。以网捕获,以乙醚深度麻醉致死。整体浸泡于10%福尔马林溶液中,观察前用自来水浸泡半日,解剖观察并取相关材料,进行石蜡切片,片厚4~6 μm,H-E 染色,观察并拍照。
为明确小肠肠绒毛的三维形态,选取一段十二指肠降部肠管,采取半连续石蜡切片,即隔片取一片进行贴片,片厚5 μm,H-E染色观察,针对某一部位或某一肠绒毛连续追踪观察。
2 结果
对器官位置的描述涉及到一定的方位术语。棕果蝠的头、颈、躯干以及四肢的相互位置关系类似于人。为了方便描述,并便于与其他动物进行比较,本文采用更加普适的方位术语,即比较解剖学的方位术语,比如距头部近者为前侧,距尾端近者为后侧,距躯干脐侧正中假想线近者为腹侧,距后背正中假想线近者为背侧。
2.1 胃
棕果蝠胃体积相对较大,伸缩性较小,空腹时仍然保持大而扁的囊状,壁薄。胃的外形和位置与人的相似。贲门位于小弯偏左侧,幽门在右后方。棕果蝠的胃形态比较特殊,如图1a所示。依据Kamiya and Pirlot[8]的划分方法,本研究将棕果蝠的胃划分为贲门部、胃底部、胃体部和幽门部4部分。
(1)贲门部。当食管穿过膈而进入腹腔后,向腹后方向延伸长约1.1 cm,并且管径逐渐变大,呈漏斗状开口于胃的小弯,将这一部分划分为胃的贲门部,有些作者常将这部分称作贲门漏斗或贲门前庭。
(2)胃底部。在贲门部右侧向前背侧突出的盲端圆顶为胃底部,外表面与贲门部有浅沟为界。
(3)胃体。胃体为贲门部漏斗所正对的部分,其与胃底部之间的界限不易确定,本研究沿用Kamiya的分界法,即以贲门部右缘的延长线划分,如图1a所示。
(4)幽门部。幽门部是由胃体开始变细的部分开始,在棕果蝠甚发达,形成总长约5 cm的袢状管道,折叠成“U”形,因此可分为幽门部降段和升段,分别长约2.3 cm。升段末端距贲门漏斗很近,然后折转向右,横行约1 cm,此时外表出现细的横向浅沟,为幽门的外在标志。其内表面有环形低矮的黏膜皱褶,即幽门括约肌,大弯和小弯处的皱褶高矮相似,属于对称性幽门括约肌。
通过组织切片观察,棕果蝠胃黏膜全部为腺体。其中胃体部、胃底部、贲门部均为胃底腺,幽门部的降段为胃底腺;幽门部升段为幽门腺。未发现无腺部或贲门腺。与大鼠胃底腺区相比,棕果蝠腺层相对较薄,每个小腺分为腺颈、腺体和腺底。腺体较长,含大量壁细胞,主细胞较少,腺底较短。幽门腺为蟠曲的管状腺,含有弱嗜碱性的黏性分泌物。
2.2 小肠
小肠始于幽门,可分为十二指肠、空-回肠两段。
2.2.1 十二指肠
十二指肠形态和位置类似于人类,可分为前水平部、前曲、降部、后曲和升部,整个十二指肠长约10 cm,如图1b。其中前水平部相对较长,约1.5 cm,具有发达的肠绒毛和浅短的肠隐窝,目测其肠绒毛呈舌状或叶片状。降部最长,约2.5 cm,沿腹腔右侧肝的腹侧面延伸到右肾的腹侧。升部较长,约1.84 cm,在其后缘可见十二指肠-结肠韧带,为菲薄的双层浆膜褶,但韧带的另一侧并没有与结肠相连,而是连于前肠系膜上。十二指肠末端形成十二指肠-空肠曲,长约0.8 cm。
对十二指肠降部进行半连续切片观察,针对某一肠绒毛连续追踪,同一肠绒毛可在20余片切片中同时出现,换算为长度,则可证明棕果蝠十二指肠降部肠绒毛为与肠管方向一致的纵向板状,长度至少在0.2 mm以上,并且众多板状绒毛排列为书页状,如图2所示。组织学观察未见十二指肠腺分布,特别是在幽门部位。
2.2.2 空-回肠
空-回肠细而长,长度在个体间变化较大,长约35~48 cm,形成十余个肠袢,由空肠系膜连于系膜根。具有发达的肠绒毛,空肠段的肠腺比大鼠的显著浅短,在某些部位,可见黏膜隆起,内含许多管状肠腺,形成集中分布的小范围隆起,在此称之为腺斑。切开肠腔,肉眼可见空-回肠黏膜形成纵向书页状肠绒毛,每片肠绒毛长约2~3 mm,每十多片肠绒毛集成丘状隆起,长宽约2~3 mm。到空-回肠末段,肠绒毛变矮,变稀少。
2.3 大肠
大肠与小肠在外观上不易区分。由于蝙蝠不存在盲肠,且大肠直径与小肠相近,不具有肠袋和纵肌带,回肠和大肠间也无明显界线,因此小肠与大肠之间的界线不易确定。为此,本研究首先确定直肠的位置和范围,然后根据肠系膜的长短确定降结肠、横结肠和升结肠,如图1c所示。对于升结肠上游的部分,通过组织切片、肠黏膜形态以及动静脉分布3方面的特征综合分析判断。
(1)组织学切片观察表明,从升结肠起始端上溯约8 cm的范围,其肠黏膜已经呈现大肠特征,即肠绒毛消失,肠腺密集出现。
(2)这一小段肠管的起始端也正是肠系膜前动脉正中干支末梢分布的范围。
(3)是从肠黏膜面上可见一条细长的纵向黏膜皱褶出现,并向后延伸,肉眼可见肠绒毛消失。
由此可知,从升结肠起始端上溯8 cm的肠管具有大肠的特征,在此将这段8 cm的肠管命名为前结肠,因此棕果蝠的大肠可分为前结肠、升结肠、横结肠、降结肠和直肠,如图1c所示。前结肠的范围为从升结肠起始端逆向追溯8 cm,肠系膜较长,活动范围较大,与空-回肠不易区分,黏膜面具有一条细线状纵行皱褶,肠绒毛消失。前结肠起始部正对肠系膜前动脉中轴支。升结肠位于右腹部,肠系膜较短,位置较固定,长度约1.4 cm,由后走向前。横结肠为升结肠与降结肠之间的过渡,长度约0.8 cm,系膜较短,位置较固定,由右走至左腹部。降结肠长约1.6 cm,后端直抵直肠,在某些个体可出现明显的乙状结肠。直肠位置固定,几无系膜,长约1.1 cm,呈前后细中间略膨大的梭状,内表面有数条纵行的皱褶。
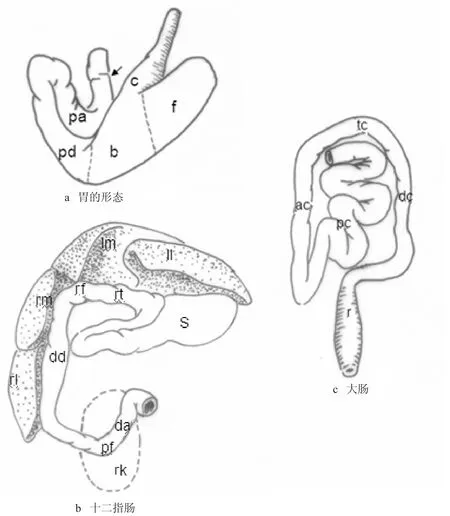
图1 棕果蝠胃、肠划分示意

图2 十二指肠肠绒毛半连续横切片
3 讨论
3.1 胃结构
本观察结果表明,棕果蝠的胃结构比较特殊,具体表现在以下几个方面。
(1)在贲门部,从外观上,食管的腹腔段相对比例显著增大,管径迅速增粗,形成漏斗状,在与胃小弯相接处达到最大,有关这一结构的名称,称为贲门漏斗或贲门前庭[8-9]。也有一些种类不具有这一结构,比如南美的绒假吸血蝠和南美大耳蝠[10]。
(2)形成外观上壶腹形状的原因有多种,报道最多的是胃腔向食管端扩展,同时胃的腺性黏膜也衬在食管端内壁。有些种类胃腔并没有实质性地向食管拓展,而是管壁的环形平滑肌显著增厚形成,如双色蹄蝠[8]。食管与胃相接处在解剖学上和组织学上并不始终一致。本实验在棕果蝠所见的即属于第一种情形,在贲门前庭作切片观察见到典型的胃底腺,是食果蝠类胃的特征之一。因此笔者将外观上的食管末段膨大部视作胃的一部分是正当的。但在棕果蝠很难从外观上断定食管与胃的确切交界处,需要采用组织切片方法作追踪观察,留待后续研究。
3.2 胃的分部
关于胃的分部,Kamiya and Pirlot提出食果蝠类可划分为3部分,即贲门漏斗、胃底盲囊和胃体[8],可能分别相当于本文的贲门部、胃底部和幽门部。该作者对于贲门漏斗所投射的区域可能并不明确。但是该作者在其同篇论文中对胃各部进行组织学取材时却采用类似于本文所采用的胃的分部方法,因此本研究在上述3部分法的基础上作出修正,将贲门漏斗前缘和后缘各线延长,其所划归的范围视为胃体部,而将原胃体部改称幽门部(图1a)。考察已报道的各类蝙蝠胃的黏膜分区[9],可知本实验观察到的幽门腺的范围相对来说是最大的,相对于食虫类、食花蜜类和食肉类及食鱼类的都要大,说明食果类的幽门腺是发达的。根据Forman[11]的观点,从食性上讲,按食虫-食蜜-食果的顺序,胃的结构是渐趋复杂的,幽门腺是由小变大的,而作为幽门腺的补偿的十二指肠腺,则是渐趋消失,在本实验中即未发现十二指肠腺。在本实验的观察中注意到无论贲门括约肌还是幽门括约肌,都是非常不明显的,与食虫类和食鱼类差别显著。有作者认为,果蝠在采食时先以咀嚼榨取果汁并吞咽之,剩余果渣则吐出口外[8],所以进入胃的为液态的果汁,本文见到的棕果蝠贲门括约肌和幽门括约肌的纤弱可能与此有关。与食虫类相比,本实验还发现棕果蝠的胃壁薄,胃黏膜层薄,皱褶很少,扩张性差,是否都与吞食果汁这一习性有关,值得探讨。
3.3 肠道
从肠道方面比较,无论是长度还是复杂性,棕果蝠都较食虫类强,主要体现在以下几个方面。
(1)肠管长度。本实验观察到棕果蝠空-回肠及结肠长约60 cm,与同等体重的食虫类蝙蝠相比长出2.5倍(本实验室资料)。
(2)结肠的发达程度。Makanya[12]对一种典型的食虫类(马铁菊头蝠)的观察发现,其结肠甚不发达,几乎可以忽略,大肠部分仅为直肠。即便是韦氏颈囊果蝠,Makanya[13]测量发现,小肠约占整个肠管长度的94%,结肠占4%,直结肠占2%。在本实验中,小肠与结肠之间的界限划分是一个棘手的问题,由于蝙蝠不存在盲肠、阑尾,且除直肠外,其他所有肠管均呈现出一致的粗细和外观,即使在某些个例出现肠管粗细不均的现象,其他例却不恒定出现。以往学者也都遇到相同的问题,比如Makanya et al[12-13]判断小肠与结肠一方面是根据粗细,另一方面根据是否出现纵向的黏膜皱褶以及肠黏膜的形态结构变化而确定。本实验中,采用多重标准来综合判断,即肠绒毛是否消失、肉眼观黏膜面是否出现细线状长的纵向皱褶,以及肠系膜内空肠脉管的分支特征加以判断,结果表明这3重指标多数情况下是一致的,取得了较为满意的结果。本实验在棕果蝠发现其具有较为完备的大肠结构,虽无盲肠和阑尾,但在升结肠上游尚有8 cm左右的前结肠,有的个体甚至具有乙状结肠。在适应飞翔而尽量简化身体结构的生存进化中,棕果蝠仍然保有相当比例的结肠,反映出大肠消化对棕果蝠可能具有重要意义。
3.4 小肠绒毛形态
小肠绒毛形态是当前国内动物解剖界的一个研究热点。刘玉堂等发现,禽小肠绒毛的三维立体形状是高板状的,与肠道方向一致,众多板状肠绒毛形成书页状结构[14]。本实验采用半连续石蜡切片观察了十二指肠降部肠绒毛的形态变化,结果清楚表明,棕果蝠小肠绒毛为纵向板状,众多板状绒毛平行排列,呈书页状。对图2中箭头所指绒毛,可连续向后追至第20片,可见其长度至少在200 μm以上。对剪开的空肠肠管观察其黏膜面,亦可肉眼观察到成片的纵向绒毛板群,它们十余片集为一群,长宽约2~3 mm,周边以矮短的板状绒毛形成浅沟,与其他绒毛板群为邻。棕果蝠小肠绒毛的特点与Makanya在马铁菊头蝠[12]报道的有显著差别,后者的肠绒毛除小肠前端和后端小部分外,主要肠段的肠绒毛都是横向板状绒毛,每片绒毛几乎环绕肠管内表面一周,形如垫片,而小肠前端和后段的肠黏膜则形成小的蜂窝状小房,状如牛羊网胃黏膜的缩微版。2001年Makanya[13]在韦氏颈囊果蝠观察到其小肠前1/5具有长的、分支的、相互吻合的肠绒毛,向后绒毛转化为指状的、相互分离的肠绒毛。由此说明,翼手目动物的肠绒毛形态具多样性,应当引起足够的关注。
