格木寄生线虫中国新记录种
2019-11-27王宏洪蔡润得黄少彬
王宏洪 蔡润得 黄少彬


摘要 在对广东省主要珍贵树种上的线虫进行调查时从格木Erythrophleum fordii上分离到一种隐皮孢囊线虫,经形态特征观察和测量数据分析,将其鉴定为卡勒萨隐皮孢囊线虫Cryphodera kalesari。其诊断特征为:雌虫椭圆形至近球形,头部具有1个唇盘和2~3个唇环,口针长33.6~37.8 μm,阴门唇突出,阴门与肛门之间区域凹陷,肛阴距为42~72 μm;2龄幼虫头部具1个明显的唇盘和3个唇后环纹,口针长24.8~29.5 μm,口针基部球前缘凹陷,侧区3条侧线,尾长圆锥形,长38~53.5 μm,尾末端细圆,透明尾长17.1~25.1 μm,侧尾腺孔位于肛门后2~5环;雄虫未发现。本研究首次获得了卡勒萨隐皮孢囊线虫的rDNA(LSU D2D3和ITS)序列,为此线虫的鉴定提供了可靠的分子数据。本研究还分析了卡勒萨隐皮孢囊线虫与本属其他种类的系统进化关系。卡勒萨隐皮孢囊线虫为中国的地理新记录种。格木为隐皮孢囊线虫的新寄主。
关键词 格木; 卡勒萨隐皮孢囊线虫; 形态学; 分子数据; 中国新记录
中图分类号: S 432.45 文献标识码: A DOI: 10.16688/j.zwbh.2019137
Abstract A species of cystoid nematode parasitizing Erythrophleum fordii was isolated during the survey of plant nematodes associated with precious trees in Guangdong province. Detailed morphological study confirmed the cystoid nematode as Cryphodera kalesari. The nematode was characterized by oval to near round females with 2-3 lip annuli and one distinct labial disc, a stylet of 33.6-37.8 μm in length, a protruding vulval lip, concave vulva-anus profile, a vulva-anus distance of 42-72 μm. Second-stage juveniles possessed 3 lip annuli and one labial disc, a stylet of 24.8-29.5 μm in length with concave knobs and 3 lines at the lateral field, an elongate-conoid tail, 38-53.5 μm long, with a narrow rounded tip, hyaline tail region of 17.1-25.1 μm long, phasmids with a lens-like structure located 2-5 annuli posterior to the anus. Male was absent. LSU rDNA D2D3 segment and rDNA-ITS fragment were amplified and sequenced, which provided molecular information for rapid and accurate detection of the cystoid nematode in the future. Moreover, the phylogenetic relationship of C.kalesari and other Cryphodera species was analyzed. C.kalesari was reported for the first time from China. Erythrophleum fordii is a new reported host plant of C.kalesari.
Key words Erythrophleum fordii; Cryphodera kalesari; morphology; molecular data; first record in China
格木Erythrophleum fordii Oliv.,別名铁木、斗登风,豆科格木属常绿乔木,生于山地密林或疏林中。广东、广西、浙江、福建、台湾等地为其天然分布区[1]。格木为国产著名硬木之一[1],被列为国家II级重点保护野生植物[2]。
隐皮孢囊属Cryphodera线虫是一类不形成孢囊(non-cyst-forming or cystoid)的异皮科Heteroderidae线虫[3]。全世界已报道7个已知种和6个未定种[4-6]。中国曾从泰国输华的柏木中截获布林克曼隐皮孢囊线虫 C.brinkmani Karssen & Van Aelst, 1999[7-8]。而国内仅报道过一种寄生苎麻的隐皮孢囊线虫——中华隐皮孢囊线虫C.sinensis Zhuo, Wang, Ye, Peng & Liao, 2014[5]。
作者在对广东省主要珍贵树种植物线虫进行调查时从格木根系上及根围土壤中采集分离到一种隐皮孢囊线虫。通过形态特征观察、测量和拍照,以及详细的形态描述,将其鉴定到种的水平。此外,我们还对该线虫的LSU rDNA D2D3区和rDNA-ITS序列进行了扩增和测序,并分析了隐皮孢囊线虫的系统发育关系。
1 材料与方法
1.1 样品采集
用内径10 cm的不锈钢土壤取样管(宏光仪器)采集广东省广州市从化区流溪河国家森林公园格木根围土壤和根系,取样深度约为20 cm,每个样品约500 g,装入自封袋,编号CH06,记录采样时间、采样地点和寄主等信息。
1.2 线虫分离、标本制作、形态测量和拍照
用60目不锈钢筛网(上虞仪器)筛出土壤中细根。在体视镜下用镊子检查细根和植物根系,从根上剥离白色雌虫(图1)。用贝曼漏斗法分离土壤中的线虫,时间为24 h。在体视显微镜下,将土壤中分离到的隐皮孢囊线虫2龄幼虫挑入清水中,加入等体积煮沸的8%甲醛溶液,虫体热杀死后至少固定24 h;用甘油酒精法对幼虫进行脱水,然后将幼虫挑入纯甘油,石蜡封片[9]。利用科研级尼康显微镜(ECLIPSE 90i,Nikon,日本)进行线虫形态测量和拍照[10]。
1.3 DNA抽提、扩增和测序
用蛋白酶K法抽提单条幼虫DNA,具体步骤参照Mundo-Ocampo等[11]。LSU rDNA D2D3区的扩增引物为D2A(5′-ACA AGT ACC GTG AGG GAA AGT-3′)和D3B(5′-TCG GAA GGA ACC AGC TAC TA-3′)[12],rDNA-ITS区扩增引物为18S(5′-TTG ATT ACG TCC CTG CCC TTT-3′)和26S(5′-TTT CAC TCG CCG TTA CTA AGG-3′)[13]。PCR反应体系参照DNA聚合酶说明书(Ex Taq,TaKaRa,日本)。PCR扩增程序、电泳检测和测序参照王宏洪等[10]。用DNAstar Lasergene 7.0软件的SeqMan程序对测序结果进行拼接和编辑,将最终序列上传到GenBank数据库,获取序列登录号。
1.4 系统进化分析
从GenBank数据库分别下载隐皮孢囊线虫属有关种的28S rDNA D2D3和rDNA-ITS序列。序列比对、核酸替代模型选择、系统进化树的构建、系统进化树的打开和编辑参考王宏洪等[10]。外組的选择参照Subbotin等[6]。基于rDNA 28S D2-D3序列构建的隐皮孢囊线虫系统进化树,外群为Scutellonema brachyurum (Steiner,1938) Andrassy, 1958 (JX472046);基于rDNA-ITS序列构建的系统进化树,外群为Hoplolaimus columbus Sher, 1963(DQ309584)。
2 结果与分析
2.1 形态鉴定结果
形态测计值见表1和表2,表中符号采用De Man公式。
雌虫形态特征:虫体椭圆形至近球形,环纹清晰,颈部向背部或腹部弯曲(图1d~e;图2A)。头部具一个明显唇盘和2~3个唇环(图2G~I)。口针长33.6~37.8 μm(图2G~H)。中食道球卵圆形,长16.3~26.3 μm,宽12.5~24.9 μm(图2B)。基部食道腺紧邻中食道球,小且不清晰(图2B)。排泄孔位于颈部之后(图2B)。阴门裂长39.4~59 μm(图2E~F)。阴门唇突出(图2A、2D)。肛阴距为42~72 μm(图2C~F)。阴门与肛门之间区域略凹陷或明显凹陷(图2C~D),具明显线纹(图2E~F)。
2龄幼虫形态特征:热杀死后,虫体直或略向腹面弯区(图3a)。侧区不明显,3条侧线(图3e)。头部具一个明显的唇盘和3个唇后环纹(图3c~d)。口针长24.8~29.5 μm,口针基部球前缘略凹陷至明显凹陷(图3c~d)。背食道腺开口至口针基部球距离3~4.5 μm(图3c~d)。食道腺充满整个体腔,长34.4~60.5 μm(图3a~b)。排泄孔至体前端距离82.4~101.5 μm(图3b)。半月体紧邻排泄孔之后(图3f)。生殖原基小,卵圆形,至尾末端距离占体长的58%~67%(图3g)。尾长圆锥形,长38~53.5 μm,尾末端细圆(图3h~i)。侧尾腺孔棱镜状,位于肛门后2~5环(图3j)。透明尾长17.1~25.1 μm(图3h~i)。
雄虫:未发现。
寄主:格木Erythrophleum fordii Oliv.。
经形态特征和测计值比对,采自广东从化格木上的隐皮孢囊线虫群体为卡勒萨隐皮孢囊线虫C.kalesari Bajaj, Walia, Dabur & Bhatti, 1989。与原始描述[14]相比,卡勒萨隐皮孢囊线虫中国群体雌虫体长偏大(336.8~481.4 μm vs 272~353 μm),口针略短(33.6~37.8 μm vs 37~42 μm),阴门裂更长(39.4~59 μm vs 35 μm),其余形态特征基本一致。卡勒萨隐皮孢囊线虫中国群体与原始描述的差异可能与地理环境有关。因此,该群体鉴定为卡勒萨隐皮孢囊线虫。
卡勒萨隐皮孢囊线虫与中华隐皮孢囊线虫形态最接近。与中华隐皮孢囊线虫相比,卡勒萨隐皮孢囊线虫雌虫口针更长(33.6~37.8 μm vs 28~35.4 μm),肛阴距更长(42~72 μm vs 29.5~35.8 μm),2龄幼虫口针更短(24.8~29.5 μm vs 28~31 μm),尾末端有尖突(vs 细圆),尾和透明尾更短(38~53.5 μm vs 52~65 μm;17.1~25.1 μm vs 24.5~35 μm)。此外,卡勒萨隐皮孢囊线虫缺乏雄虫。
2.2 分子序列分析
经扩增和测序,获得卡勒萨隐皮孢囊线虫中国群体LSU rDNA D2D3区片段长度为699 bp,GenBank的登录号为MK541650。BLAST结果显示,此群体与Cryphodera sp. B(MF425671)的序列相似性最高,为93.7%;与布林克曼隐皮孢囊线虫(KY512575)和中华隐皮孢囊线虫(JX566455)的序列相似性次之,为92.3%。
扩增得到的卡勒萨隐皮孢囊线虫中国群体rDNA-ITS片段长度为1 122 bp,GenBank的登录号为MK541651。BLAST结果显示,此群体与中华隐皮孢囊线虫(JX566457-JX566459)的序列相似性最高,为76.6%~76.8%。
2.3 系统进化分析
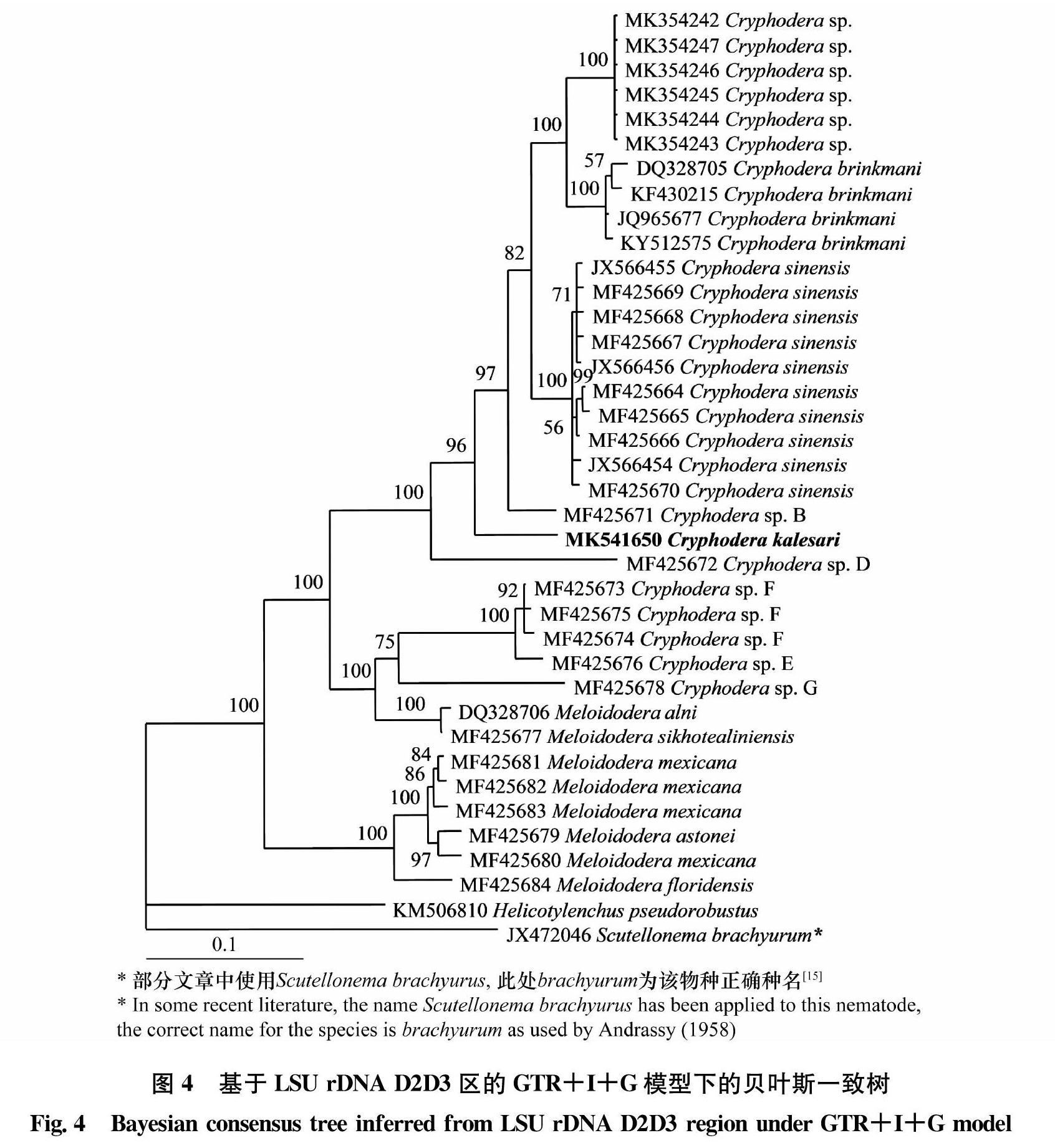
rDNA 28S D2-D3区序列比对数据集包含38条序列,比对后的长度为684 bp,其中保守位点422个,简约信息位点185个。图4为基于rDNA 28S D2 D3 序列在GTR+I+G模型下的贝叶斯一致树,卡勒萨隐皮孢囊线虫与Cryphodera sp. B、中华隐皮孢囊线虫、布林克曼隐皮孢囊线虫和Cryphodera sp. 聚为一个高度支持的分支(PP=96),并处于分支的基部。rDNA-ITS区序列比对数据集包含41条序列,比对后的长度为1 183 bp,其中保守位点266个,简约信息位点846个。图5为基于rDNA-ITS序列在TVM+I+G模型下的贝叶斯一致树,卡勒萨隐皮孢囊线虫与Cryphodera sp. B、Cryphodera sp.和布林克曼隐皮孢囊线虫聚为一个中度支持的分支(PP=86),卡勒萨隐皮孢囊线虫与其他隐皮孢囊线虫种类的系统进化关系不清楚。
3 结论与讨论
本研究将采自广东省广州市从化区良口镇流溪河国家森林公园格木的隐皮孢囊线虫鉴定为卡勒萨隐皮孢囊线虫,该线虫为中国新记录种。本文首次获得了卡勒萨隐皮孢囊线虫的28S rDNA D2D3区和rDNA-ITS序列,为今后隐皮孢囊线虫的种类鉴定提供了分子依据。
Bajaj等1989年最早在印度哈里亚纳邦卡勒萨森林的羽萼木Colebrookea oppositifolia Smith根上发现卡勒萨隐皮孢囊线虫[14]。此线虫广泛分布于位于印度哈里亚纳邦和喜马偕尔邦的西瓦里克山脉的天然林中[16],分布区的气候类型为热带季风气候,主要特点是全年暖热,降水明显分旱、雨两季[17]。本次报道的卡勒薩隐皮孢囊线虫分布区的气候类型是热带季风气候,气候温和,雨量充沛[18]。
本文明确了格木为卡勒萨隐皮孢囊线虫的新寄主。受侵染的格木根系表面无明显症状,雌虫嵌入根系,突出的尾部通常黏附土壤颗粒,不容易冲洗干净。格木是我国重要的珍贵树种,线虫病害是格木苗期的主要病害,但线虫种类未知[19]。本研究首次报道了侵染格木根系的一种隐皮孢囊线虫,其对格木生长的危害程度仍需进一步研究。
目前,已报道的7个隐皮孢囊属有效种中有3个发现于亚洲,分别是卡勒萨隐皮孢囊线虫C.kalesari(印度和中国),布林克曼隐皮孢囊线虫C.brinkmani(日本和泰国)和中华隐皮孢囊线虫C.sinensis(中国和越南)[6]。Nguyen等从越南采集了大量隐皮孢囊线虫种群,通过比对分析2龄幼虫的形态测量数据和rDNA-ITS序列数据,认为其中至少存在5个种类[20]。Subbotin等重新分析了来自越南的样品,推测其中有7个种类,除了分布最广泛的中华隐皮孢囊线虫,另外还有6个潜在新种[6]。近年来,我们在华南地区各省份也陆续发现一些隐皮孢囊线虫种群,由于仅发现2龄幼虫,缺乏雌虫形态数据,未能鉴定到种。因此,中国南部和越南可能是隐皮孢囊线虫的起源和多样性中心,说明隐皮孢囊线虫在这个区域正在进行相对快速率的物种形成[6]。
参考文献
[1] 中国科学院中国植物志编辑委员会.中国植物志(第39卷),豆科(一)[M].北京:科学出版社,1988:117.
[2] 于永福.中国野生植物保护工作的里程碑——《国家重点保护野生植物名录(第一批)》出台[J].植物杂志,1999,5(3):3-11.
[3] SIDDIQI M R.Tylenchida,parasites of plants and insects [M].London:CABI,2000.
[4] VOVLAS N,TRISCIUZZI N,TROCCOLI A,et al.Integrative diagnosis and parasitic habits of Cryphodera brinkmani a non-cyst forming heteroderid nematode intercepted on Japanese white pine bonsai trees imported into Italy [J].European Journal of Plant Pathology,2013,135(4):717-726.
[5] ZHUO Kan,WANG Honghong,YE Weimin,et al.Cryphodera sinensis n.sp.(Nematoda:Heteroderidae),a non-cyst-forming parasitic nematode from the root of ramie Boehmeria nivea in China [J].Journal of Helminthology,2014,88(4):468-480.
[6] SUBBOTIN S A,AKANWARI J,NGUYEN C N,et al.Molecular characterisation and phylogenetic relationships of cystoid nematodes of the family Heteroderidae (Nematoda:Tylenchida) [J].Nematology,2017,19(9):1065-1081.
[7] 龙海,齐小峰,谢泳桂,等.泰国输华真柏中一种隐皮孢囊线虫的鉴定[J].植物检疫,2017,31(3):29-32.
[8] KARSSEN G,AELST A.Description of Cryphodera brinkmani n.sp.(Nematoda:Heteroderidae),a parasite of Pinus thunbergii Parlatore from Japan,including a key to the species of the genus Cryphodera Colbran,1966 [J]. Nematology, 1999, 1(2):121-130.
[9] 冯志新.植物线虫学[M].北京:中国农业出版社,2001.
[10] WANG Honghong,ZHUO Kan,YE Weimin,et al.Morphological and molecular characterisation of Pratylenchus parazeae n.sp.(Nematoda:Pratylenchidae) parasitizing sugarcane in China[J].European Journal of Plant Pathology,2015,143(1):173-191.
[11] MUNDO-OCAMPO M,TROCCOLI A,SUBBOTIN S A,et al.Synonymy of Afenestrata with Heterodera supported by phylogenetics with molecular and morphological characterisation of H.koreana comb.n.and H.orientalis comb.n.(Tylenchida:Heteroderidae) [J].Nematology,2008,10(5):611-632.
[12] SUBBOTIN S A,STURHAN D,CHIZHOV V N,et al.Phylogenetic analysis of Tylenchida Thorne,1949 as inferred from D2 and D3 expansion fragments of the 28S rRNA gene sequences [J].Nematology,2006,8(3):455-474.
[13] VRAIN T C,WAKARCHUK D A,LEVESQUE A C,et al.Intraspecific rDNA restriction fragment length polymorphism in the Xiphinema americanum group [J].Fundamental and Applied Nematology,1992,15(6):563-573.
[14] BAJAJ H K,WALIA R K,DABUR K R,et al.Cryphodera kalesari,a new heteroderid nematode species from Haryana,India [J].Systematic Parasitology,1989,14(2):113-116.
[15] ANDRSSY I.Hoplolaimus tylenchiformis Daday,1905 (syn.H.coronatus Cobb,1923) und die gattungen der unterfamilie Hoplolaiminae Filipjev,1936 [J]. Nematologica,1958(3):44-56.
[16] BAJAJ H K,WALIA R K.Host response to Cryphodera kalesari (Heteroderidae:Nematoda)[J].Nematologia Mediterranea,2000,28(2):197-199.
[17] 張兆明.浅析热带季风气候的差异性[J].地理教育,2010(10):26-27.
[18] 吴章文.流溪河国家森林公园旅游气候研究[J].中南林学院学报,1995(10):67-74.
[19] 李胜强,许建新,陈波,等.珍稀植物格木的研究进展[J].广东林业科技,2008,24(6):61-64.
[20] NGUYEN C N,STURHAN D,SUBBOTIN S A.Studies on the occurrence and diversity of Heteroderidae and Meloidogynidae (Nematoda:Tylenchida) in natural forests of Vietnam [J].Russian Journal of Nematology,2011,19(2):159-172.
(责任编辑: 田 喆)
