油茶象种群密度及空间格局
2019-11-27李志文赵瑞李有志
李志文 赵瑞 李有志

摘要 油茶象Curculio chinensis Chevrolat是我國特有木本油料树种——油茶Camellia spp.的专性蛀果害虫,常导致其大量落果。本文旨在明确油茶象幼体(卵+幼虫)种群动态、空间格局及其关键影响因子,以便为调查取样、长期监测和科学防控等提供理论依据。2017年在湖南常宁普通油茶林,2016年-2017年在湖南益阳小果油茶林于雌虫产卵季节进行了调查,解剖收集的果实并记录其所含幼体数。用6种聚集度指标(m*/m、Iδ、C、I、Ca、k)和Taylor幂法则、Iwao回归模型分析油茶象幼体的空间格局,并探讨其关键影响因子。调查和分析表明,雌虫于6月上、中旬开始产卵,幼体种群密度呈单峰变化,2016年和2017年高峰期分别在7月25日和7月18日。小果油茶林2016年、2017年高峰期种群密度分别为17.95±1.53和16.27±1.06幼体/样株,高于普通油茶林,分别是普通油茶林种群密度(8.16±1.23幼体/样株)的2.20倍和1.99倍,差异极显著(P<0.001)。聚集度指标分析显示,油茶象在普通油茶林均呈聚集分布,在小果油茶林呈聚集分布或随机分布。回归模型表明,该虫在两种油茶林均呈聚集分布。Blackith种群聚集均数λ>2,表明油茶象幼体聚集是由亲代雌虫本身的产卵行为与寄主果实特征综合作用的结果。给出了最适理论抽样数公式并绘制了3种抽样精度下最适抽样数随种群密度变化的曲线图。综上,初步明确了油茶象幼体在普通油茶林和小果油茶林的空间分布格局和种群动态,并探讨了其关键影响因子。试验结果可为该虫调查取样、长期监测和科学防控等提供理论依据。
关键词 油茶象; 普通油茶; 小果油茶; 种群密度; 空间格局; 最适抽样数
中图分类号: Q 968 文献标识码: A DOI: 10.16688/j.zwbh.2018379
Abstract The camellia weevil, Curculio chinensis Chevrolat, is an important pest attacking fruits of Camellia spp., an endemic genus in China, and causes fruit drop. To provide theoretical basis for survey sampling, long-term monitoring and scientific prevention and control of the weevil pest, its spatial distribution pattern and population fluctuation were investigated. During the egg-laying season of the weevil, systematic investigations were conducted on the Camellia oleifera farm in Changning city, Hunan province in 2017 and C.meiocarpa farm in Yiyang city, Hunan province in 2016 and 2017, respectively. The fruits collected were dissected and the weevil immatures contained in them were recorded; then the spatial distribution pattern of the immatures was analyzed using 6 aggregation indices (m*/m,Iδ, C,I,Ca and k) and 2 regression models (Taylors Power Law and Iwaos Patchiness Regression), and finally key factors affecting spatial pattern were identified. The egg-laying of the weevil females initiated in early to mid-June. The population density of the immatures showed a single peak change and the highest density was recorded on 18th July and 25th July. The density on C.meiocarpa farm was obviously higher than that on C.oleifera farm in the same season; the peak density of the former was 17.95±1.53 immatures per tree in 2016 and 16.27±1.06 immatures per tree in 2017, 2.20 and 1.99 times higher than that of the latter (8.16±1.23 immatures per tree) in 2017, respectively. The analysis of aggregation indices showed that it was aggregated on C.oleifera farm for each sampling of the immatures, but random or aggregated distribution on C.meiocarpa farm. The regression models indicated that the weevil immatures spatial distribution pattern was clustered on each farm. It was proposed that the combined action of the oviposition behavior of the parent females and its host fruit traits led to the aggregation distribution pattern of the weevil immatures, as the Blackith λ value was more than 2. The formula of optimal sample size and the curve charts of optimum sample size changing with population density of the weevil immature were presented. In conclusion, the spatial pattern, population density and fluctuation of the weevil immatures were clarified, and the key influencing factors were also discussed. The experimental results can provide a theoretical basis for survey sampling, long-term monitoring and scientific prevention and control of the weevil pest.
Key words Curculio chinensis; Camellia oleifera; Camellia meiocarpa; population density; spatial pattern; optimum sample size
油茶象Curculio chinensis Chevrolat属鞘翅目Coleoptera象甲科Curculionidae,是我国特有木本食用油料树种——油茶Camellia spp.专性蛀果害虫[1-2],被国家林业局列入“全国林业危险性有害生物名单”(国家林业局防止外来林业有害生物管理办公室2003年发布)之中,在我国大多数油茶产区均有分布。普通油茶Camellia oleifera、小果油茶C.meiocarpa、腾冲红花油茶C.reticulate和广宁红花油茶C.semiserrata均受其害[1-5],也可为害茶C.sinensis和刺锥栗Castanopsis hystrix[6]。
成虫取食以长喙上的口器咬住油茶果实,身体在果面上往返旋转,喙钻穿果皮后伸达种子内部,吸食种子汁液或咬食果仁。卵通过喙钻蛀的孔道即刺孔产入寄主果实果仁内。幼虫蛀食果仁,在单个果实内发育,发育到4~5龄常导致寄主果实脱落,落果油茶象寄生率1.5%~75.3%[7]。最严重的落果率可达96.6%[8],严重影响油茶产量和品质[9]。
目前,有关油茶象的研究主要涉及为害程度[3-4,8,10]、生物学[11-14]、生物防治[15-17]及其与寄主植物协同进化[7,18-19]等方面。对其种群密度及季节动态的研究未见详细报道,而这是研究农林害虫的基础,尤其是害虫管理方面如为害程度监测、损失程度估计和确定防治措施等均有赖于此[20]。昆虫的空间分布格局是其行为与环境因子综合作用的结果,了解昆虫的空间分布型及其关键影响因子是保证调查取样可靠性的关键[21],有助于理解害虫的种群特性,是进行科学防控的基础[22-25]。害虫种群动态及其空间分布格局通常具有地域性[20]。赵丹阳等于7月上旬对广东韶关油茶象幼虫的空间分布型进行了初步调查[26]。本研究在我国油茶大省湖南于油茶象产卵季节对普通油茶和小果油茶进行了系统调查,旨在明确该地区油茶象幼体种群密度季节动态及其空间分布格局,并探讨其关键影响因子,试图从行为学的角度探讨亲代雌虫产卵行为对子代幼体分布格局的影响,有利于从整体上把握油茶象的种群特性,期望为油茶象调查取样、长期监测及科学防控等提供科学依据。
1 材料与方法
1.1 研究区概况
调查的普通油茶林位于湖南省常宁市罗桥镇。该地位于常宁市南偏东约15 km处(26°17′N,112°26′E,海拔116 m),丘陵地貌,属亚热带季风性湿润气候,四季分明,雨量充沛。年均气温约18.1℃,日照1 600 h,降水量约1 436 mm,无霜期近300 d。试验林面积约7 hm2。树龄为60~70年,高度为2.5~5.5 m。调查的小果油茶林位于湖南省益阳市赫山区。该地位于益阳市东南约5 km处(28°29′N,112°26′E,海拔64 m),地势较为平坦,属亚热带大陆性季风湿润气候,光照充足,雨量充沛。年均气温16.1~16.9℃,日照1 348~1 772 h,降雨量1 230~1 700 mm,无霜期263~276 d。试验林面积约3 hm2,树龄为50~60 年,高度为2.5~5.5 m。两样地多年均未进行人工抚育等管理措施,林内郁闭度较高,林下植被以白茅Imperata cylindrical为主,间有芒萁Dicranopteris dichotoma、刺莓Rubus taiwanianus和柃木Eurya sp. 等植物分布。
1.2 试验方法
2017年于普通油茶林采用“Z”形取样法,选择结果实较多的样树(因需多次取样,下同)25株。2016年和2017年于小果油茶林分别选择结果实较多的样树21株和22株。在油茶象产卵季节大约每隔10 d调查一次,每季调查6~7次(具体日期见图1)。每样树按东南西北中、上下层确定10个取样点,每点随机采摘果实2个,共采摘20个果实。采摘的果实按样树编号分别装袋,带回实验室解剖检查,记录每果幼体(卵+幼虫)数,解剖工作2~3 d内完成。
1.3 数据统计分析
采用Excel 2010、SPSS 13.0和Origin 9.0进行数据分析与作图。日均气温参考当地气象资料,为最高气温和最低气温平均值。用独立样本t检验(independent-samples t test)分析种群密度高峰期普通油茶林和小果油茶林油茶象幼体密度差异性。
1.3.1 分布型测定
计算各样地每次调查油茶象幼体平均值m,方差S2,平均拥挤度m*=m+S2/m-1,Beall扩散系数C=S2/m,David和Moore的丛生指数I=S2/m-1,Morisita扩散指数Iδ=n∑(fx2-N)/(N(N-1))≈m*/(m-1/n)(N为总虫数,n为抽样数),Waters负二项分布k值k=m2/(S2-m),Lloyd聚块性指数m*/m,Cassie和Kunor的聚集度指数Ca=(S2-m)/m2等参数,依照其判别标准判定空间分布型[27-28]。
1.3.2 聚集原因檢验
利用Taylor(1961)幂法则S2=amb,即lg(S2)=lg(a)+blg(m)和Iwao(1971)的m*-m线性回归模型m*=α+βm分析油茶象幼体的空间分布格局和种群结构。用Blackith种群聚集均数λ=mγ/(2k)检验聚集原因[27],式中,m为平均种群密度,k为负二项分布的k值,γ为自由度为2k时0.5水平下的χ2值。
1.3.3 确定最适抽样数
依据Taylor幂法则最适理论抽样公式Nopt=tα/2D2amb-2和Iwao回归方程最适理论抽样公式Nopt=tα/2D2α+1m+(β-1)[29-32] 分别计算最适理论抽样数。式中,D为相对误差(允许误差的百分比,即D=d/m,d为允许误差),a和b为Taylor幂法则回归系数,α和β为Iwao回归方程参数,m为平均种群密度,t为置信度。
2 结果与分析
2.1 油茶象种群密度及其季节动态
小果油茶上6月14日未见幼虫,只见少量卵,为1.44±0.27(mean±SE,下同)卵/样株,即0.07卵/果(图1)。此后每次调查卵和幼虫均有出现。幼体种群密度具明显高峰期,普通油茶林(图1Ⅰ)和2016年小果油茶林(图1Ⅱ)均于7月25日达最大值,分别为8.16±1.23幼体/样株和17.95±1.53幼体/样株。2017年小果油茶林油茶象幼体种群密度高峰期稍提前,7月18日即达峰值(图1Ⅲ,16.27±1.06幼体/样株)。高峰期后,种群密度均急剧下降,但直至8月31日(图1Ⅱ,6.33±0.62幼体/样株)和9月3日(图1Ⅰ,3.28±0.59幼体/样株)还保持一定水平。相同季节小果油茶林幼体种群密度大于普通油茶林,高峰期前者密度分别为后者密度的2.20倍和1.99倍,差异极显著(t=-5.042,P<0.001;t=-4.991,P<0.001)。
2.2 油茶象种群空间分布格局
对普通油茶林7次调查中(表1),聚块性指数、扩散指数和扩散系数(m*/m,Iδ,C)均>1,丛生指数和聚集度指数(I,Ca)均>0,负二项分布k值,0
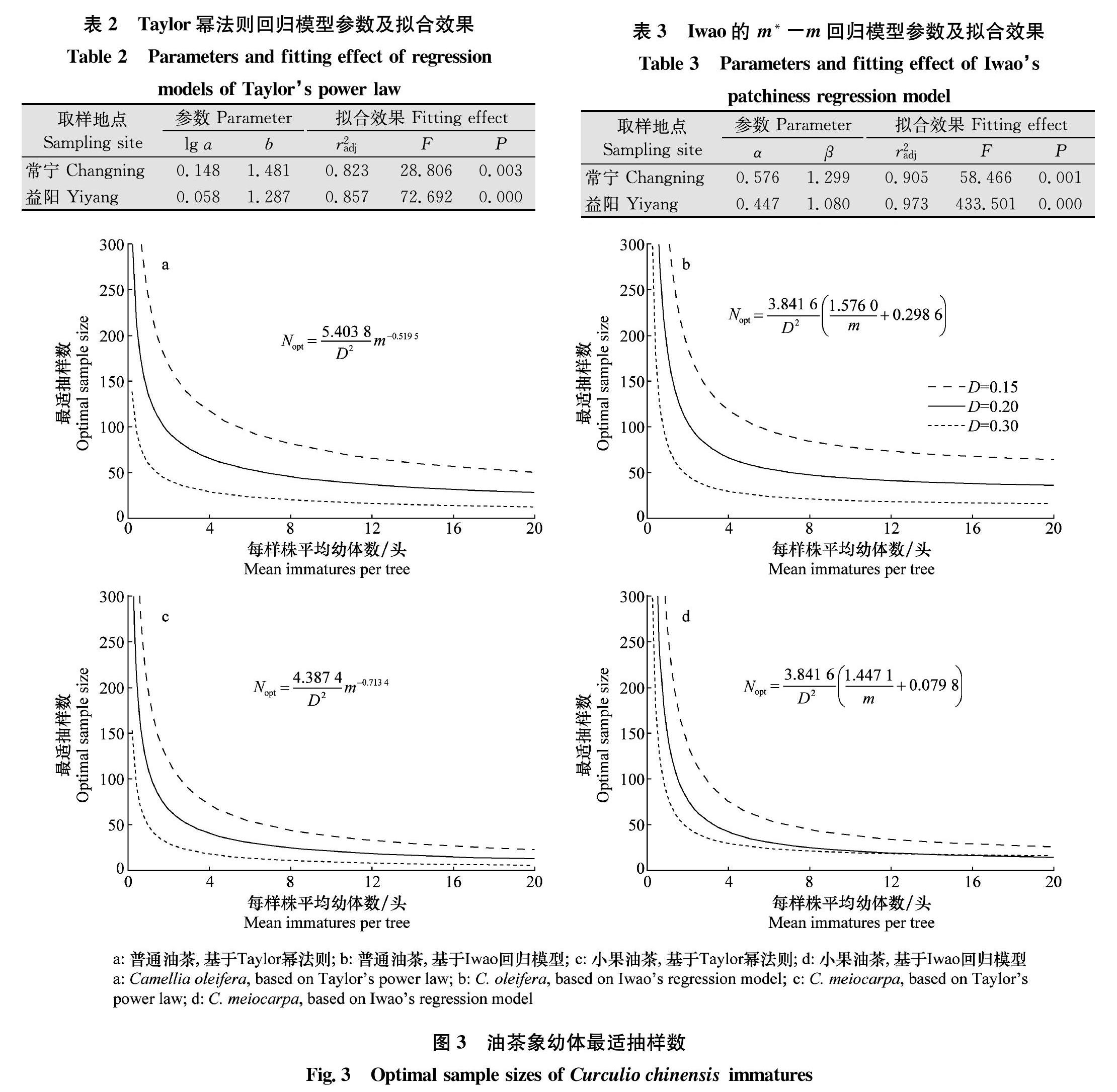
根据Taylor幂法则,对lg S2和lg m进行回归分析,结果显示普通油茶林和小果油茶林二者均呈显著线性正相关(图2a)(r2adj=0.823,P=0.003;r2adj=0.857,P<0.001),回归参数见表2。由于lg(a)>0,b>1,说明种群在一切密度下均是聚集的,且随着油茶象种群密度增大,聚集度增高。根据Iwao回归模型,对m*和m进行线性回归分析,表明二者线性关系极显著(图2b)(r2adj=0.905,P=0.001;r2adj=0.973,P<0.001),回归参数如表3。α>0,β>1,表明油茶象幼体间相互吸引,分布的基本成分为个体群,且呈聚集分布。Blackith种群聚集均数λ>2(表1),表明聚集是由油茶象本身的行为习性引起,或是由其生活习性与环境因子综合作用的结果。但2017年6月14日对小果油茶的调查表明,λ=1.360<2,说明此次聚集可能是某些环境因素导致的。
2.3 最适理论抽样数
用Taylor和 Iwao回归系数,给出了油茶象最适抽样数理论公式,并分别绘制了不同误差允许水平(D=0.15,0.20和0.30;t=1.96,95%置信概率)最适理论抽样数随种群密度变化的曲线(图3)。比较表明,两种回归模型计算出的理论抽样数变化趋势基本一致。种群密度越低,抽样数越大。误差允许水平越高,抽样数越小。Iwao模型较Taylor模型線性拟合优度更高(前者r2adj≥0.905,后者r2adj≤0.857,表2和表3)。相同误差允许水平,通过Iwao模型计算出的最适理论抽样数略高。
3 结论与讨论
本研究中整个调查季节(6月14日至9月3日)均能调查到卵,说明油茶象产卵期长,至少110 d以上,与前人报道[11]结果(150 d)一致。赵丹阳7月上旬在韶关5样地调查的幼虫密度为3.88~12.75头/样株(即0.06~0.21头/果),低于本研究同期(7月5日)普通油茶林油茶象幼体种群密度(4.65±0.45幼体/样株,即0.23幼体/果),符合预期,因为前者种群密度不包括卵,且油茶林树龄为10年左右。油茶象具有卵单产的行为习性,正常条件下,雌虫产卵于寄主果实果仁内,幼虫孵化后在单个果实内发育,果内幼体(卵+幼虫)为同一雌虫或不同雌虫多次产卵的累积效应[18-19]。这说明油茶象幼体空间分布格局和种群动态主要取决于亲代雌虫的产卵行为和繁殖力,因此影响亲代雌虫产卵行为和繁殖力的因子也必然会对子代分布格局和种群动态产生作用。一般而言,昆虫的产卵行为和繁殖力除与其自身生理状态有关外,主要受温度、寄主植物品种、寄主植物物候及寄主果实特征(如果实大小、果皮厚度、木质化程度及种皮硬化情况等)等的影响。
3.1 油茶象种群密度的关键影响因子
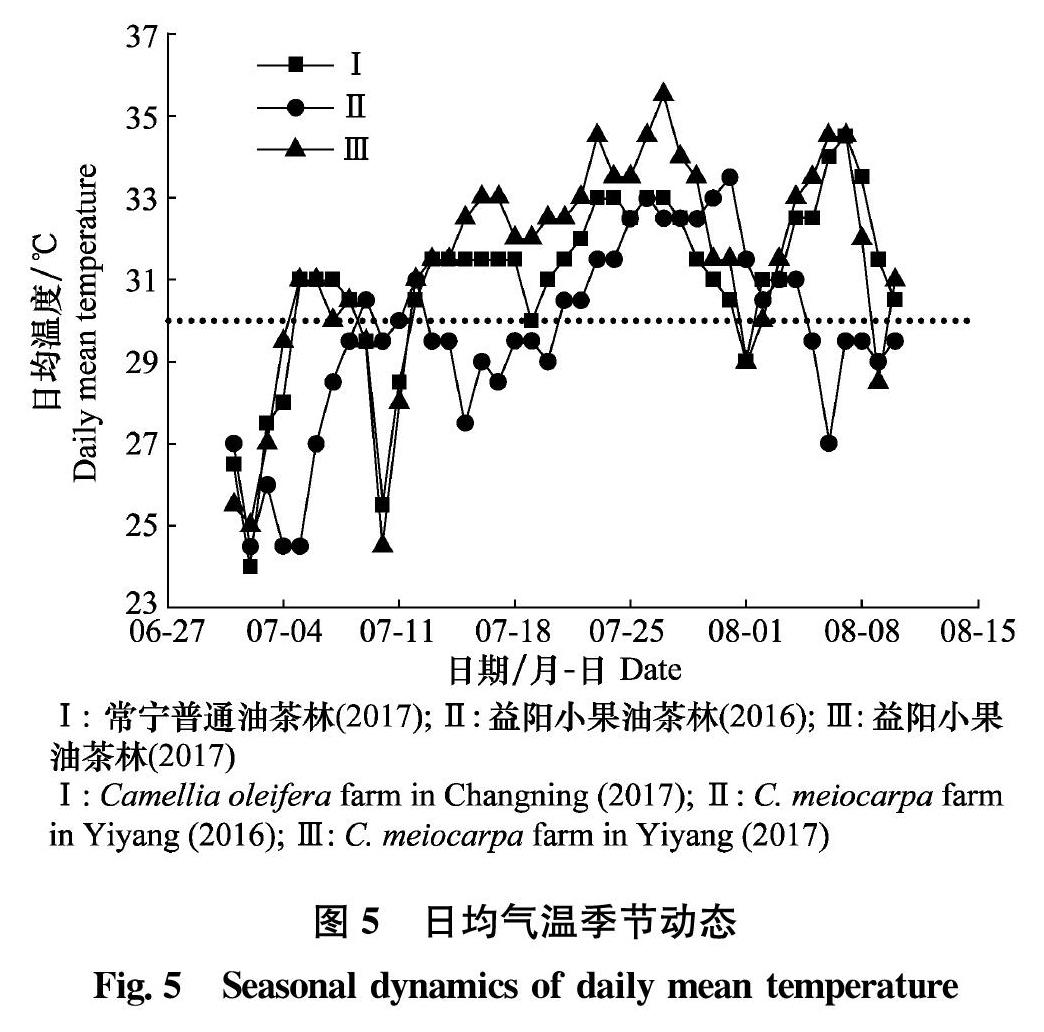
据赵丹阳等[5]报道,油茶象不在果径<13 mm的果实上产卵。 6-8 月是油茶果实及种子迅速生长期[33]。本研究中,6月初随机采摘小果油茶果实100个,仅3个果实具刺孔,解剖无虫卵。6月14日调查均有卵粒(1.44±0.27卵/样株,即0.07粒卵/果)。据报道卵期10~15 d[11],说明产卵起始于6月上、中旬。此时平均果径为(14.74±0.09)mm,种内营养物质已开始聚集(图4b),为雌虫提供了相对嫩枝嫩叶更为丰富而优质的营养,从补充营养的角度来说这是合理的。随着寄主果实继续发育,幼体种群密度急剧上升,于7月中、下旬达最大值后显著下降(图1),这可能有以下几个方面的原因。首先,象蛀果实大量脱落。据报道,小果油茶林象蛀果实7月中、下旬开始大量脱落[7],普通油茶林中象蛀果实脱落延迟,于7月下旬至8月初开始(未发表数据)。二是连续高温导致雌虫产卵量下降。马玲等报道连续数日最高气温>35℃会导致油茶象成虫大量死亡[34]。进一步研究表明,日均温度高于29℃显著影响油茶象雌虫寿命,32℃恒温较29℃恒温雌虫日均产卵量急剧减少(未发表数据)。Zahiri等亦报道,不利高温(31.5℃恒温较29℃恒温)会导致紫苜蓿叶象Hypera postica雌虫寿命和日均产卵量呈断崖式下跌[35]。湖南地区7月中旬至8月初通常高温干旱,2016年益阳赫山7月20日至8月3日的日均气温≥30.5℃,2017年两样地7月12日始日均气温≥30.5℃,且一直持续至8月10日(图5)(其中只有2~3 d日均气温低于30.5℃)。2017年不利高温较2016年来临较早,可能导致了雌虫产卵量提前降低和雌虫过早死亡,因而幼体高峰期较2016年稍微提前(图1)。第三,寄主种皮硬化。寄主种皮于7月中、下旬开始硬化,引起雌虫摄食、产卵困难(图4d),进而导致其快速死亡。种皮变硬后,卵只能产于种皮外,幼虫孵化后因不能钻入种内摄食,仅取食果皮很难完成正常发育,多在低龄期(1~2龄)死亡(图4e)。以上几个方面综合作用,影响亲代雌虫营养和寿命,进而影响其繁殖力,也影响后代幼虫的存活,加上虫蛀果实提前脱落,必然导致油茶象幼体种群密度呈单峰型变化。
3.2 油茶象空间格局的关键影响因子
油茶象幼体在不同斑块(样树或果枝)的密度是亲代雌虫在该斑块持续产卵的累计效应。卵通过取食孔产入寄主果实果仁内,产卵、取食行为密切联系[19]。震落法表明,成虫多聚集在果实密度较大处。根据最优觅食理论[36],捕食者在优质资源斑块(样树或果枝)停留时间较劣质斑块长。对油茶象雌虫而言,优先选择高密度果枝(样树)或长时间停留,降低被捕食风险的同时,可有效缩短寄主果实搜索时间,提高摄食效率。成虫可在果面反复钻孔取食,单果刺孔数最高可达20多个,因而可以认为优质斑块食物资源是无限的。但产卵情况截然不同,由于小果实对幼虫发育具限制作用[18],为增加子代适合度,亲代雌虫具优先选择大果实和卵单产的行为习性[19]。这提示雌虫对果内卵和幼虫可能存在某种识别机制。由此可见,油茶象雌虫优先选择优质资源斑块并长时间停留,满足食物资源需求的同时,优先将卵聚集到优质资源斑块。这有利于成虫释放繁殖力[37]。随着卵的不断聚集,雌虫搜寻未产卵果实的时间逐渐增长,当达到某一临界值时就有可能转移到其他资源斑块。根据此假说,当种群密度低时,幼体呈聚集分布,随着种群密度升高,聚集度逐渐增强,即使所有果实均被寄生,幼体亦呈聚集分布,因为寄主果实本身疏密不一。本试验中,Taylor幂法则和Iwao回归模型表明,油茶象幼体间相互吸引,分布的基本成分为个体群,种群在一切密度下均呈聚集分布,且具密度依赖性,即m越高,聚集度越高,符合假说预期。赵丹阳等[26]在韶关的调查亦具类似结果。6种聚集度指标分析表明,每次调查普通油茶林均呈聚集分布,小果油茶林共进行了13次调查,其中5次调查(2016年7月25日、7月31日及2017年7月6日至7月27日的3次调查)综合判定呈随机分布,这可能是受虫蛀果实脱落的影响,象虫蛀食果实脱落前各次调查呈聚集分布。聚集原因分析显示,总共20次调查中,19次调查Blackith种群聚集均数λ>2(表1),表明油茶象幼体聚集是由亲代雌虫本身的产卵行为与寄主果实特征综合作用的结果。
3.3 油茶象监测及防控
本研究调查的油茶林为50~70 年老林,多年未进行人工抚育管理,缺乏正常垦覆等措施对油茶象入土虫态的控制,从而导致种群密度较正常管理油茶林偏高。多次调查需求致使取样偏向选择结果较多的样树,可能导致种群密度进一步升高。尽管如此,由于所选油茶林缺少人为干扰,所得结果更能真实、客观地反映油茶象种群自然属性,有利于正确理解油茶象与其寄主植物的相互作用和机制。目前有关油茶象为害程度的报道已有很多[2-4, 8,10,38-39],但由于调查方法和日期不同或难以考证,因而生产实践中很难有效借鉴。一般而言,由于受雌虫生理状态、气候条件、寄主果实特征以及虫蛀果实脱落等多种因素的影响[7,18-19,34,40],油茶象幼体林间种群密度随时间往往不断变化。本文明确了其单峰动态模型,并探讨了温度、寄主种皮硬化和虫蛀果实脱落等关键因子的影响,据此建议油茶象长期监测中调查适期应选择在幼体种群密度高峰期,因为其他时段均有较高的低估风险。高峰期一般在7月中、下旬,各地可依据其关键影响因子的不同或作用强度进行适当调整。在确定取样精度的基础上,参考图3确定样本大小。油茶象幼体的聚集分布格局既反映了亲代雌蟲的产卵行为,又预示了后继虫态的分布规律,一定程度上可为油茶象调查取样、采取针对性和差异化的防控措施提供科学依据。
参考文献
[1] 庄瑞林.中国油茶[M].第2版.北京:中国林业出版社,2008:366.
[2] 蔡守平,何学友,李志真,等.油茶象危害油茶果实的初步研究[J].福建林业科技,2011,38(2):14-16.
[3] 伍建榕,穆丽娇,林梅,等.滇西地区红花油茶主要病虫害种类调查[J].中国森林病虫,2012,31(1):22-26.
[4] 谢胤.腾冲红花油茶古树病虫、寄生植物危害调查及保护建议[J].林业调查规划,2012,36(6):77-80.
[5] 赵丹阳,秦长生,徐金柱,等.油茶象甲成虫对油茶寄主选择性研究[J].中国农学通报,2015,31(17):100-104.
[6] 张汉鹄,谭济才.中国茶树害虫及其无公害治理[M].合肥:安徽科学技术出版社,2004:389.
[7] 李志文,孙寒松.湖南益阳小果油茶林小果油茶、油茶象和啮齿动物的相互作用[J].昆虫学报,2016,59(10):1123-1132.
[8] 蒋三俊.油茶象鼻虫的防治[J].特种经济动植物,2009,12(8):54.
[9] 李苗苗,张威,吕军美,等.茶籽象为害对油茶果产量和茶油品质的影响[J].植物保护,2016,42(5):65-68.
[10] 李密,周刚,彭争光,等.湖南油茶害虫风险性评估及危险性等级划分[J].中国农学通报,2014,30(19):277-283.
[11] 周石涓.油茶象的生物学及其防治[J].昆虫学报,1981,24(1):48-52.
[12] 徐磊,潘涌智,黄阳洋.油茶象的潜在分布区预测[J].福建林业科技,2011,38(4):55-58.
[13] 李志文,何立红,夏姣,等.油茶象幼虫虫龄的划分[J].昆虫学报,2015,58(2):181-189.
[14] 赵丹阳,秦长生,徐金柱,等.油茶象甲形态特征及生物学特性研究[J].环境昆虫学报,2015,37(3):681-684.
[15] 邓小军,周国英,刘君昂,等.N+注入选育高毒力球孢白僵菌菌株及对3种油茶害虫的毒力测定[J].中国生物防治学报,2012,28(3):341-347.
[16] 何学友,蔡守平,杜月飞,等.感染油茶象幼虫的高致病力金龟子绿僵菌菌株筛选[J].林业科学,2015,51(8):52-59.
[17] 何学友,蔡守平,詹祖仁,等.绿僵菌对油茶象幼虫林间控制效果[J].福建林业科技,2015,42(4):18-22.
[18] 李志文,何立红,马玲,等.果实大小对油茶象幼虫生长的影响[J].应用生态学报,2014,25(12):3580-3586.
[19] 李志文,何立红,杨柳君,等.小果油茶上油茶象的产卵策略[J].昆虫学报,2015,58(9):981-988.
[20] MORADI-VAJARGAH M,GOLIZADEH A, RAFIEE-DASTJERDI H, et al. Population density and spatial distribution pattern of Hypera postica (Coleoptera: Curculionidae) in Ardabil, Iran [J]. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 2011, 39(2): 42-48.
[21] BOEVE P J, WEISS M. 1998. Spatial distribution and sampling plans with fixed levels of precision for cereal aphids (Homoptera: Aphididae) infesting spring wheat [J]. Canadian Entomologist, 1998, 130: 67-77.
[22] RODEGHIERO M, BATTISTI A. Inter-tree distribution of the spruce web-spinning sawfly, Cephalcia abietis, at endemic density [J]. Agricultural and Forest Entomology, 2000,2(4): 291-296.
[23] CHO K, LEE J H, PARK J J, et al. Analysis of spatial pattern of greenhouse cucumbers using dispersion index and spatial autocorrelation [J].Applied Entomology & Zoology, 2001, 36(1): 25-32.
[24] EBER S. Bottom-up density regulation in the holly leaf-miner Phytomyza ilicis [J]. Journal of Animal Ecology, 2004, 73(5): 948-958.
[25] VINATIER F, TIXIER P, DUYCK P F, et al. Factors and mechanisms explaining spatial heterogeneity: a review of methods for insect populations [J]. Methods in Ecology & Evolution, 2011, 2(1):11-22.
[26] 赵丹阳,叶燕华,方天松,等.油茶象甲幼虫发生及空间分布型研究[J].广东农业科学,2015(12):78-81.
[27] 丁岩钦.昆虫数学生态学[M].北京:科学出版社,1980:654.
[28] 徐汝梅.昆虫种群生态学[M].北京:北京师范大学出版社,1988:409.
[29] TAYLOR L R. Aggregation, variance and the mean [J]. Nature, 1961, 189(4766): 732-735.
[30] IWAO S. A new regression method for analyzing the aggregation patter animal populations [J].Researches on Population Ecology, 1968, 10(1): 1-20.
[31] BUNTIN G D.Developing a primary sampling program [M]∥PEDIGO L P, BUNTIN G D.Handbook of Sampling Methods for Arthropods in Agriculture. Boca Raton: CRC Press, 1994:99-115.
[32] YOUNG L J, YOUNG J H. Statistical ecology: a population perspective [M].Kluwer Academic Publishers, Norwell, MA. 1998: 565.
[33] 周長富.油茶种子发育过程组分及脂类代谢相关基因表达变化研究[D].北京:中国林业科学研究院,2013.
[34] 马玲,曾爱平,李志文.油茶象生物学初步研究[M]∥王满囷,朱芬,帅移海,等.华中昆虫研究.北京:中国农业科学技术出版社,2015:132-138.
[35] ZAHIRI B, FATHIPOUR Y, KHANJANI M et al. Modeling demographic response to constant temperature in Hypera postica (Coleoptera: Curculionidae) [J]. Journal of Economic Entomology, 2010, 103(2): 292-301.
[36] CHARNOV E J. Optimal foraging: the marginal value theorem [J].Theoretical Population Biology,1976,9(2):129-136.
[37] SCHEIRS J, BRUYN L D.Integrating optimal foraging and optimal oviposition theory in plant-insect research [J]. Oikos, 2002, 96(1):187-191.
[38] 赵丹阳,秦长生,揭育泽,等.广东省油茶病虫害种类及发生动态调查[J].安徽农业科学,2012,40(29):14267-14270.
[39] 舒金平,滕莹,刘剑,等.油茶采前落果原因初步分析[J].中国植保导刊,2013,33(1):9-14.
[40] 马玲,朱桂兰,曾爱平.油茶象甲研究进展[J].湖南林业科技,2017,44(3):84-89.
(责任编辑: 田 喆)
