Toll样受体2基因敲除通过myd88依赖的p38MAPK减轻高脂饮食诱导的肥胖小鼠脂肪组织氧化应激
2019-11-27宋冰王茹张浩强
宋冰 王茹 张浩强
(1锦州医科大学附属第一医院,辽宁 锦州 121000;2锦州医科大学研究生学院)
随着患病率逐年增高,肥胖已经成为世界性难题〔1,2〕,它可以引起胰岛素抵抗、2型糖尿病、心脏病、终末期肾病甚至是某些癌症等一系列疾病〔3〕。肥胖机体脂肪组织存在低水平的炎症反应〔4〕及氧化应激〔5〕在肥胖发生发展中的作用已经被广泛研究,近年来高游离脂肪酸(FFA)能够激活Toll样受体(TLR)2越来越引起研究者的关注,FFA激活TLR2后可层层激活包括myd88依赖的炎症通路而引起下游炎症因子的表达〔6〕,而包括肿瘤坏死因子(TNF)-α在内的炎症因子表达和氧化应激反应密切〔7〕,本实验利用高脂饮食诱导肥胖小鼠,并检测肥胖小鼠脂肪组织炎症因子以及氧化应激状态的改变,然后再利用高脂饮食喂养的TLR2基因敲除的小鼠探究TLR2基因敲除对肥胖小鼠脂肪组织炎症因子表达和氧化应激状态的影响。
1 材料和方法
1.1实验动物分组 SPF级健康4周龄雄性C57bl/6小鼠和TLR2基因敲除小鼠各24只,随机分为正常对照组(NC组)(8只),高脂饮食组(HFD组)(16只),TLR2基因敲除肥胖组(TK组)和TLR2基因敲除高脂饮食组(TH组)。NC组和TK组给予普通饮食(华阜康,北京),HFD组和TH组给予含60%脂肪的高脂饮食(华阜康,北京)。16 w后HFD组随机挑选8只体重大于NC组上限纳入肥胖组(OB组),TH组随机挑选8只体重大于TK组上限者纳入TO组(TO组)用于实验,其余剔除。各组小鼠饲养于锦州医科大学附属第一医院实验动物中心,所有动物实验符合锦州医科大学伦理委员会规定。
1.2生化指标检测 各组小鼠禁食6 h,心脏取血,检测空腹血糖(FPG)、三酰甘油(TG)、总胆固醇(TC)、高密度脂蛋白(HDL)、低密度脂蛋白(LDL)和FFA。
1.3Western印迹 动物处死后速取腹腔脂肪组织,-80℃冰箱储存备用,冰上取100 mg脂肪组织,剪碎后加裂解液裂解(RIPA∶PMSF=100∶1),聚氰基丙烯酸正丁酯(BCA)蛋白定量试剂盒(博奥森,北京)定量至2 mg/ml,聚丙烯酰胺凝胶电泳,赛默飞Marker标注蛋白位置,湿转蛋白及Marker至聚偏氟乙烯(PVDF)膜,TLR2(博奥森,北京),myd88(万类,沈阳),p38MAPK(博奥森,北京)和β-actin(ABSCI,美国)一抗4℃孵育过夜,过氧化物酶标记的二抗(ABSCI,美国)常温孵育2 h,超敏电化学发光(ECL)试剂盒(万类,沈阳)及GISt020凝胶图像分析仪成像。
1.4荧光实时定量PCR 腹腔脂肪组织总RNA采用Trizol抽提,含反转录酶的反转录试剂盒(Bioneer,韩国)反转录mRNA为cDNA,QIAGEN Fast Cycling PCR 试剂盒(Takara生物科技,大连)用于扩增:按照95℃(预变性,10 s;变性5 s),60℃(退火,15 s),70℃(拉伸,15 s)进行扩增45个循环。引物由上海生工设计并合成,白细胞介素(IL)-6:正义5′-ATGAAGTTCCTCTCTGCAAGAGACT-3′,反义5′-CACTAGGTTTGCCGAGTAGATCTC-3′;TNF-α:正义5′-TGTCTCAGCCTCTTCTCATT-3′,反义5′-AGATG-ATCTGAGTGTGAGGG-3′;GAPDH:正义5′-TTGTCAAGCTCATTTCCTGGTATG-3′,反义5′-GGATAGG-GCCTCTCTTGCTCA-3′,所得数据以GAPDH作为内参,计算IL-6和TNF-α基因的相对拷贝数,相对拷贝数用2-△△Ct表示。
1.5氧化应激检测 脂肪组织1∶20的比例加入100 mmol/L磷酸缓冲液匀浆后离心(12 000 r/min)10 min,取中层清液,BCA定量如上,调整蛋白浓度至2 mg/ml后,活性氧簇(ROS)检测试剂盒(万类,沈阳)检测活性氧含量。丙二醛(MDA)检测试剂盒和超氧化物歧化酶(SOD)检测试剂盒(建成生物研究所,南京)检测MDA含量和SOD活力。
1.6统计分析 析因设计分析交互作用,单因素方差分析组间差异,数据处理采用SPSS20.0软件。
2 结 果
2.1生化指标 与NC组相比,OB组小鼠FPG,TC,TG,LDL和FFA明显升高(P<0.01),HDL明显降低(P<0.01)。与OB组相比,TO组FPG,TC,TG,HDL,LDL和FFA均降低(P<0.01,P<0.05),且TLR2基因敲除和高脂饮食喂养之间存在交互作用(表1)。
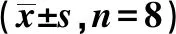
表1 各组血脂、血糖、FFA水平比较
与NC组比较:1)P<0.01,2)P<0.05;与OB组比较:3)P<0.01,4)P<0.05;与TK组比较:5)P<0.01,6)P<0.05,下表同
2.2Western印迹 与NC组相比,OB组小鼠TLR2、myd88和p38MAPK蛋白水平明显升高(P<0.01);而与OB组相比,TO组myd88和p38MAPK明显降低(P<0.01),见图1、表2。
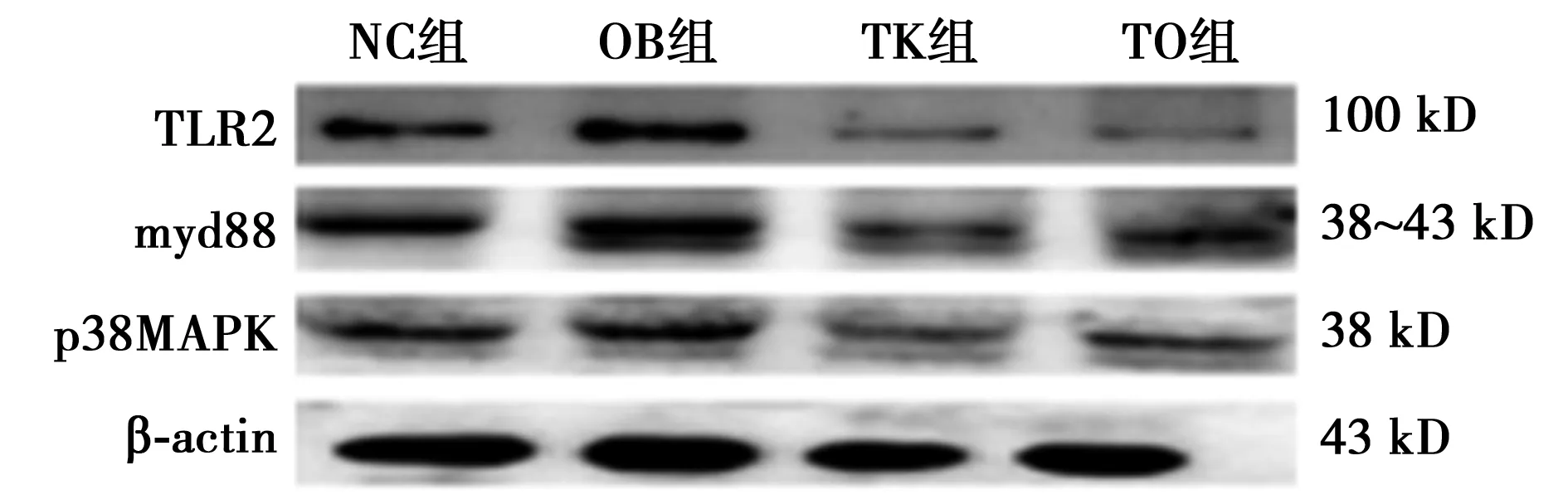
图1 TLR2,myd88,p65nfkb和p38MAPK蛋白相对表达量

组别TLR2myd88p38MAPKNC组0.22±0.030.72±0.0150.26±0.04OB组0.73±0.041)1.02±0.061)0.48±0.031)TK组-0.75±0.070.13±0.01TO组-0.86±0.113)0.17±0.013)
2.3实时荧光定量PCR 与NC组相比,OB组小鼠脂肪组织IL-6和TNF-α mRNA水平明显升高,与OB组小鼠相比,TO组小鼠脂肪组织IL-6和TNF-α mRNA水平明显降低,且TLR2基因敲除和高脂饮食喂养对其影响存在交互作用(P=0.000),见表3。
2.4氧化应激指标检测 与NC组相比,OB组小鼠脂肪组织ROS和MDA水平明显升高,SOD活力水平明显降低,与OB组小鼠相比,TO组小鼠脂肪组织ROS和MDA水平明显降低,SOD活力明显增加,见表4。
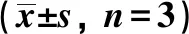
表3 各组TNF-α、IL-6 mRNA表达量比较

表4 各组脂肪组织氧化应激指标水平比较
3 讨 论
近年来超重和肥胖已经越来越成为亟待解决的问题〔1,8〕。肥胖机体中的高水平FFA可以刺激TLR2而层层激活炎症,刺激炎症因子的表达〔6〕,我们猜想,在高脂饮食诱导的肥胖小鼠体内可能存在TLR2及其下游炎症因子的高表达,myd88依赖的炎症通路是TLRs激活后引起炎症反应的重要通路〔6〕,而其下游通过p38MAPK影响炎症因子的表达〔9〕。本实验结果与之前研究认为肥胖小鼠脂肪组织IL-6和TNF-α高表达结果一致〔10〕,炎症因子的表达可使组织处于氧化应激状态〔7,11〕。所以我们猜想肥胖小鼠可以通过myd88依赖的p38MAPK途径激活下游的炎症因子并使脂肪组织处于氧化应激状态,ROS是诱导体内发生氧化应激的主要物质〔12〕,能攻击生物膜中的多不饱和脂肪酸,引发脂质过氧化作用,并因此形成脂质过氧化物,如MDA,SOD对机体的氧化与抗氧化平衡起着至关重要的作用,可清除超氧产物,可作为机体还原能力水平的检测指标。本实验结果表明高脂饮食诱导的肥胖小鼠脂肪组织处于氧化应激状态。由于肥胖小鼠脂肪组织TLR2高表达,我们猜想脂肪组织发生的炎症反应及氧化应激状态可能与TLR2激活有关,所以我们采用TLR2基因敲除小鼠并给予高脂饮食以探究TLR2基因敲除对高脂饮食诱导的肥胖小鼠脂肪组织炎症因子的表达和氧化应激状态的影响。结果显示TLR2基因敲除抑制了myd88,p38MAPK,IL-6 mRNA和TNF-α mRNA的高表达,并减轻了脂肪组织的氧化应激状态,综上所述,TLR2基因敲除可能通过myd88依赖的p38MAPK途径减轻了高脂饮食诱导的肥胖小鼠脂肪组织的氧化应激状态,但是这一结论需要进一步利用双基因甚至多基因敲除动物模型做进一步验证。
