小麦-中间偃麦草蓝粒代换系的创制与鉴定
2019-11-22叶晓斌范仁春张相岐
叶晓斌,卫 波,范仁春,张相岐
(中国科学院遗传与发育生物学研究所,植物细胞与染色体工程国家重点实验室,北京 100101)
小麦(TriticumL.)籽粒颜色一般为白色或红色,但也有一些蓝色、紫色等特殊粒色的品种和种质资源。在一粒小麦(T.monococcumL.)、二粒小麦(T.aethiopicumJakubz.)和普通小麦(T.aestivumL.)中都有蓝粒的报道[1-2]。小麦的蓝色籽粒是因为在糊粉层中有蓝色花青素积累[3]。花青素具有抗氧化、抗癌和降血糖等功效[4-5]。并且,蓝粒小麦还含有较丰富的氨基酸和微量元素等对人体健康有利的成分,因而具有较高的营养保健价值[6-7]。在小麦遗传育种中,蓝色籽粒可以作为标记性状加以利用。李振声等[8]通过普通小麦与十倍体长穗偃麦草远缘杂交方法培育了蓝单体小麦。进而,穆素梅等[9]利用蓝单体培育出了稳定遗传的小麦缺体材料。利用蓝粒标记无需进行大量的细胞学鉴定,只凭籽粒颜色就可以鉴别二体、单体或缺体材料。黄寿松等[10]利用小偃麦的蓝粒性状标记选育了雄性不育-保持系,解决了小麦核型雄性不育系保持困难的问题。Hanson等[11]以蓝粒为标记性状研究隔离距离对小麦品种间异交率的影响,确定了保证种子纯度的最小隔离距离。
小麦的一些近缘属种也有蓝粒性状,如偃麦草属(Thinopyrum=Elytrigia)、山羊草属(Aegilops)、黑麦属(Secale)等。Jan等[12]将十倍体长穗偃麦草(Elytrigiapontica=Thinopyrumponticum=Agropyronelongatum)中控制蓝粒性状的染色体4Ag(=4E)导入普通小麦后,得到了4EL/4AS易位系,并将蓝粒基因定位到4Ag染色体长臂上。后来,Zheng等[13]又将长穗偃麦草的蓝粒基因Ba1定位到4Ag染色体长臂0.71~0.80区间,并开发了与蓝粒基因连锁的分子标记和FISH探针[14]。百萨偃麦草(Thinopyrumbessarabicum=Elytrigiabessarabica)的蓝粒基因被命名为BaThb,并被定位于4Eb染色体长臂的着丝粒与FL0.52之间的区域[15]。Love 和Suneson[16]以及袁文业和孙善澄[2]曾报道在普通小麦与中间偃麦草(Et.intermedia)的杂交后代中也出现了蓝粒材料,但未见进一步研究。来自于栽培一粒小麦(T.monococcum,2n=2x=14, AA)的蓝粒基因被命名为Ba2,并已被定位于4AL的着丝粒附近[17]。
为给蓝粒小麦品种的选育提供新的种质资源,并为中间偃麦草蓝粒基因的遗传学研究奠定基础,本研究利用八倍体小偃麦中5(来自普通小麦与中间偃麦草杂交)与普通小麦中国春的缺-四体系列材料杂交和回交,在后代中选育出两份蓝粒材料。通过细胞学和分子标记鉴定,确定了两份蓝粒小麦材料的染色体组成和控制蓝粒性状染色体的来源染色体组和同源群。
1 材料与方法
1.1 材 料
八倍体小偃麦中5(2n=8x=56,AABBDDXX)来自于普通小麦(2n =6x=42, AABBDD)与中间偃麦草(2n=6x=42, EeEeEeEeStSt或EeEeEbEbStSt)的杂交后代,由原黑龙江省农科院孙善澄研究员培育[18],种子由东北师范大学郝水院士惠赠。一套普通小麦中国春(CS)的缺体-四体(NT)系列材料的种子由英国John Innes Centre的John Snape教授惠赠。二倍体长穗偃麦草(2n=2x=14,EeEe)的种子由美国密苏里大学的Perry Gustafson教授惠赠。假鹅观草[Pseudoroegneriastipifolia(Nevski) Love,2n=2x=14,StSt]的种子由中国农业科学院的李立会研究员惠赠。中国春-二倍体长穗偃麦草[Et.elongata(Host) Nevski]的双二倍体CS/THE(2n=8x=56, AABBDDEeEe)种子由中国科学院遗传与发育生物学研究所的王道文研究员惠赠。百萨偃麦草[Et.bessarabica(Savul.and Rayss) Dubovik,2n=2x=14,EbEb(= JJ)]基因组DNA由中国科学院遗传与发育生物学研究所的韩方普研究员惠赠。普通小麦中国春种子由本实验室收集保存。
1.2 方法
1.2.1 染色体制片
染色体制片参考Han等[19]的方法,略有 修改。
1.2.2 连续GISH-FISH分析
染色体的连续GISH-FISH鉴定参照Han等[19-20]的方法。GISH探针标记体系为60 μL,包括要标记的基因组DNA 6.0 μg,加重蒸水至 35.0 μL,10×Nick buffer 6.0 μL,dNTP (不含dUTP,1 mmol/L)5.0 μL,DNA polymerase I 10.0 μL,DNase 2.0 μL,Fluorescein-12-dUTP(用于中间偃麦草基因组DNA标记)或Texas Red-5-dCTP(用于二倍体长穗偃麦草、百萨偃麦草和假鹅观草基因组DNA标记)2.0 μL。混合液15 ℃金属浴标记反应2 h,然后加入480 μL ssDNA和1 mL(90%乙醇 + 10% NaAc),混匀后-20 ℃过夜。12 000 r·min-1离心30 min,收集沉淀,用70%乙醇冲洗2次,晾干,加60.0 μL 2×SSC溶解并将探针-20 ℃避光保存。
GISH杂交液体系为10 μL,包括荧光标记的偃麦草或假鹅观草探针0.5 μL,封阻DNA (中国春基因组DNA,浓度为3.5 μg·μL-1) 4.5 μL, 2×SSC 5.0 μL。选取制备好的染色体制片,放入交联仪中交联两次。将杂交液(每张染色体制片10 μL)滴加到染色体制片上的根尖酶解液区域,加塑料盖玻片,放入垫有湿润滤纸的容器内,沸水浴5 min。将水浴过的染色体制片放入垫有湿润滤纸的载玻片盒中(内有载玻片支架),55 ℃处理12 h后取出,2×SSC溶液洗脱盖玻片。滴加10 μL DAPI复染液,盖上24×50 mm盖玻片,荧光显微镜下观察和照相。选取有较好有丝分裂中期相的染色体制片,用70%乙醇洗脱DAPI及荧光标记后用于进一步的FISH分析。
FISH检测所使用的探针为来自于黑麦(SecalecerealeL.)的重复序列pSc119.2和来自于粗山羊草(AegilopstauschiiCoss.)的重复序列pAs1。探针pSc119.2用Fuorescein-12-dUTP标记为绿色,探针pAs1 用Texas Red-5-dCTP标记为红色。FISH探针标记和杂交程序参考Han等[20]的方法。
1.2.3 SNP标记分析
本研究所使用的SNP标记为中国科学院遗传发育所的娄海娟博士[21]通过二倍体长穗偃麦草(Et.elongata) Ee基因组与普通小麦中国春直向同源基因之间的SNP位点对比开发的Ee基因组特异SNP标记,共373对引物,分布在1E至7E染色体上。SNP标记分析采用PCR扩增方法。使用全式金公司的EASYTaq构建20 μL PCR体系,包括模板DNA 1.0 μL(100 ng·μL-1),正、反向引物各0.3 μL(0.1 μmol·L-1),dNTP混合物1.6 μL(dNTPs 2.5 mmol·L-1),Taq酶0.2 μL(EASYTaqDNA Polymerase 1.0 U),10×EASYTaqbuffer (200 mmol·L-1Tris-HCl,200 mmol·L-1KCl,100 mmol·L-1(NH4)2SO4)2.0 μL,LC Green荧光剂(Idaho) 1.0 μL,ddH2O 13.6 μL。程序为: 94 ℃ 预变性5 min;94 ℃ 变性30 s,56 ℃ 退火20 s,72 ℃ 延伸10 s,35个循环;72 ℃ 延伸10 min。结果分析采用高分辨率熔解曲线(HRM)方法。高分辨率熔解曲线检测仪器为Light Scanner(Idaho, LightScanner 96)。
1.2.4 SSR标记分析
SSR标记来自小麦。使用GenStar Mix构建20 μL PCR体系。程序为:95 ℃变性5 min; 95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,35个循环;72 ℃延伸10 min。PCR扩增产物用8%聚丙烯酰胺凝胶电泳分离。
2 结果与分析
2.1 蓝粒小偃麦的选育
以八倍体小偃麦中5作母本,中国春缺-四体系列材料作父本进行杂交获得杂种F1,用缺-四体亲本作母本回交一次后连续自交至BC1F4代。每代选取植株形态偏向中国春的类型混收并繁育下一代。结果分别在中5×N4BT4A和中5×N7BT7D两个杂交组合的后代植株中发现了蓝色籽粒(图1)。分别收获蓝粒种子并自交繁殖一代。田间观察显示,蓝粒株系与白粒姊妹系在株穗型上并无明显差别。选取蓝粒性状不发生分离单株Zh5-a2-1(来自中5×N4BT4A组合)和Zh5-c13-2(来自中5×N7BT7D组合)的种子进行染色体组成分析。

a:中国春;b:中国春缺-四体系N4BT4A;c:中国春缺-四体系N7BT7D;d:中5;e:蓝粒株系Zh5-a2-1;f:蓝粒株系Zh5-c13-2。
a:Chinese Spring; b:Chinese Spring NT line N4BT4A; c:Chinese Spring NT line N7BT7D; d:Zhong 5; e:Blue-grained line Zh5-a2-1; f:Blue-grained line Zh5-c13-2.
图1 两份蓝粒小麦及其亲本和中国春的种子
Fig.1 Seeds of the two blue-grained wheat lines and their parents and Chinese Spring
2.2 蓝粒小偃麦的染色体组成
2.2.1 蓝粒小偃麦的染色体组成
利用中间偃麦草基因组DNA作探针的GISH分析显示,蓝粒系Zh5-a2-1和Zh5-c13-2的根尖细胞染色体数均为42条,包括40条小麦染色体和2条中间偃麦草染色体(图2a, c),即一对中间偃麦草的染色体代换了一对小麦的染色体。进一步利用pSc119.2和pAs1作探针对同一染色体制片进行FISH分析,通过与中国春标准核型[22]比对,发现两个蓝粒系Zh5-a2-1和 Zh5-c13-2中被代换的一对小麦染色体分别为4B和4D(图2b,d)。并且,两个蓝粒系中的一对中间偃麦草染色体有非常相似的形态大小和一致的FISH带型,都在短臂靠近着丝粒区有一pAs1探针的杂交信号(图2b和d中箭头所示)。由此推断,蓝粒系Zh5-a2-1和Zh5-c13-2分别是4B和4D染色体的二体代换系,并且二者可能代入了同一对中间偃麦草染色体。本研究也对两个蓝粒代换系的多个白粒姊妹系进行了平行GISH鉴定,结果显示在所有白粒姊妹系中均不存在中间偃麦草的染色体(图2e, f)。由此说明,这两个蓝粒代换系的蓝粒性状是由代入的一对中间偃麦草染色体决定的。
2.2.2 蓝粒代换系中中间偃麦草染色体的染色体组来源
由于尚没有中间偃麦草的标准FISH核型可供参考,因此,仅根据FISH带型尚不能判定蓝粒代换系Zh5-a2-1和Zh5-c13-2中的中间偃麦草染色体的染色体组来源和同源群归属。研究表明,中间偃麦草的染色体组构成可能为EeEeEeEeStSt[23]或EeEeEbEbStSt[24],但中5中的中间偃麦草染色体的所属染色体组还不清楚。为了明确蓝粒代换系中的中间偃麦草染色体的染色体组来源,分别利用二倍体假鹅观草(StSt)、二倍体长穗偃麦草(EeEe)和百萨偃麦草(EbEb)的基因组DNA作探针,用小麦基因组DNA做封阻,对代换系Zh5-a2-1进行了GISH鉴定,同时以中间偃麦草基因组DNA为探针的GISH作对照。结果发现,Zh5-a2-1中的一对中间偃麦草染色体只能与St基因组DNA探针杂交,而不能与Ee或Eb基因组DNA探针杂交(图3)。当用St基因组DNA探针杂交时,杂交信号分布于Zh5-a2-1的整条中间偃麦草染色体上,且信号强度与用中间偃麦草基因组DNA作探针的杂交信号相似(图3a,b)。而当用Ee或Eb基因组DNA探针杂交时,Zh5-a2-1中的一对中间偃麦草染色体则完全没有杂交信号(图3c,e),但作为对照的同一细胞用中间偃麦草基因组DNA作探针杂交时,则都能产生很强的杂交信号(图3d,f)。由此推断,蓝粒代换系Zh5-a2-1中的一对中间偃麦草染色体属于St组。由于连续GISH-FISH分析结果显示,另一个蓝粒代换系Zh5-c13-2中的一对中间偃麦草染色体与Zh5-a2-1中的相同(图2),所以,可推定Zh5-c13-2中的一对中间偃麦草染色体也属于St组。另外,用St基因组DNA作探针对中5的GISH分析结果显示,在中5中有7对中间偃麦草染色体,其中两对染色体在整条染色体上都有很强的St基因组DNA探针的原位杂交信号,推测其属于St组。而另外5对染色体只有节段性的St组DNA探针杂交信号,推测其不属于或不完全属于St组染色体(结果未在本文中显示)。由此说明中5中确实存在St组的染色体。
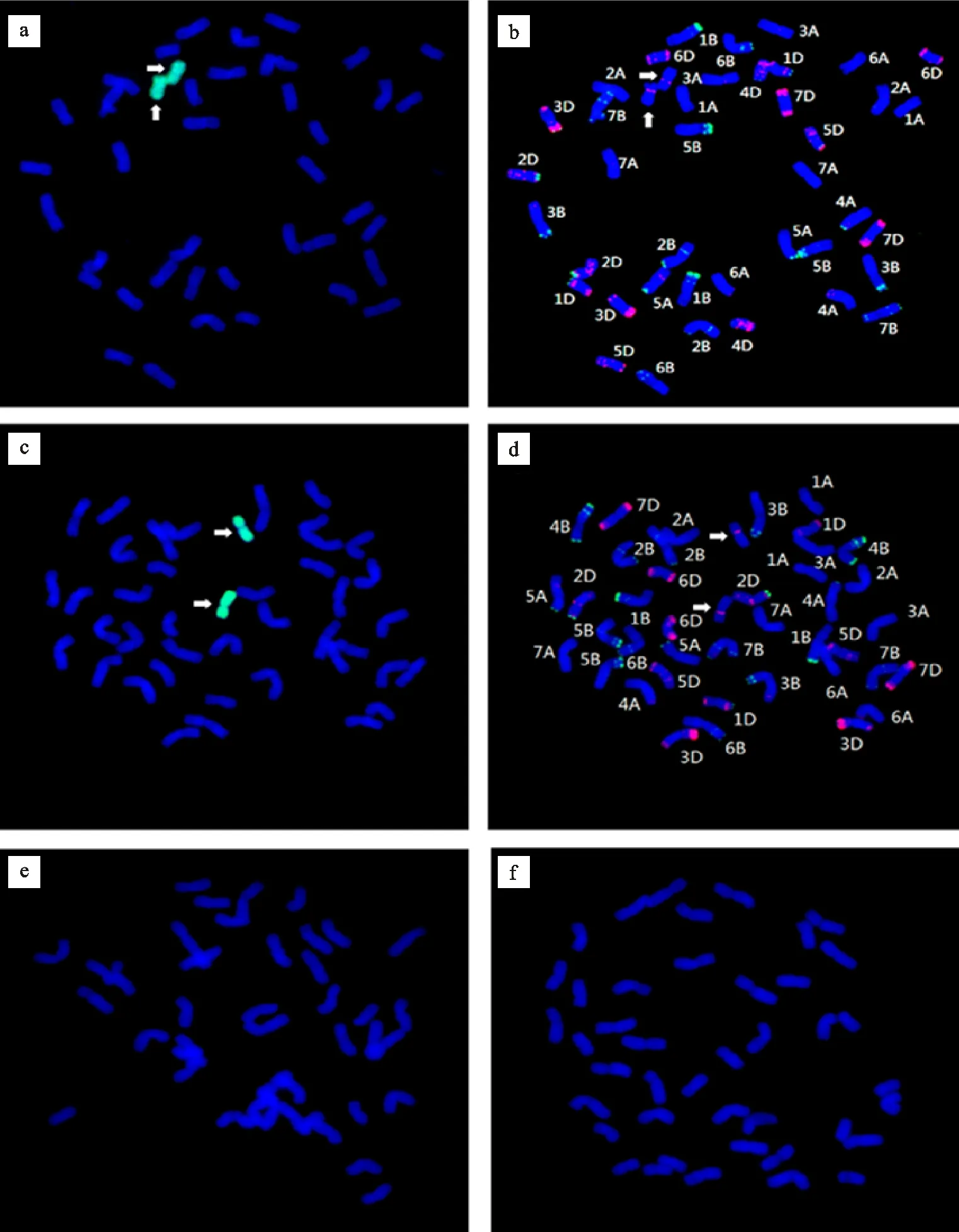
a:蓝粒代换系Zh5-a2-1染色体的GISH鉴定结果;b:与图a同一细胞染色体的FISH鉴定结果;c:蓝粒代换系Zh5-c13-2染色体的GISH鉴定结果;d:与图c同一细胞染色体的FISH鉴定结果;e:蓝粒代换系Zh5-a2-1的白粒姊妹系Zh5-a2-2染色体的GISH鉴定结果;f:蓝粒代换系Zh5-c13-2的白粒姊妹系Zh5-c13-3染色体的GISH鉴定结果。GISH探针为Fluorescein-12-dUTP标记的中间偃麦草基因组DNA;FISH探针分别为Fuorescein-12-dUTP标记的pSc119.2 (绿色) 和Texas Red-5-dCTP标记的pAs1 (红色);箭头示一对中间偃麦草染色体。
a:GISH identification of chromosomes in blue-grained substitution line Zh5-a2-1; b:FISH identification of the chromosomes in the same cell as subfigure a; c: GISH identification of chromosomes in blue-grained substitution line Zh5-c13-2; d:FISH identification of the chromosomes in the same cell as subfigure c. e:GISH identification of chromosomes in a white-grained sib line of blue-grained substitution line Zh5-a2-1; f: GISH identification of chromosomes in a white-grained sib line of blue-grained substitution line Zh5-c13-2.The Fluorescein-12-dUTP labelled genomic DNA ofEt.intermediawas used as GISH probe; The Fuorescein-12-dUTP labelled repetitive sequence ofS.cerealepSc119.2(green) and Texas Red-5-dCTP labelled repetitive sequence ofAe.tauschiipAs1(red) were used as FISH probes; The arrows indicate the pair ofEt.intermediachromosomes.
图2 小麦-中间偃麦草蓝粒代换系染色体的连续GISH-FISH鉴定结果
Fig.2Identifications of chromosomes of common wheat-Et.intermedia blue-grained substitution lines by sequential GISH-FISH
2.2.3 蓝粒代换系中的中间偃麦草染色体同源群归属
以上的细胞遗传学分析已证明Zh5-a2-1和Zh5-c13-2中被代换的小麦染色体同属第四同源群,分别为4B和4D(图2),代入的中间偃麦草染色体属于St组(图3),但其同源群归属仍不能确定。已有的研究表明,中间偃麦草和十倍体长穗偃麦草都有Ee和St染色体组,并且两者有很近的亲缘关系[25]。因此,本研究利用来自于二倍体长穗偃麦草Ee基因组的SNP标记[21],通过高分辨率溶解曲线(HRM)方法对蓝粒代换系Zh5-a2-1和Zh5-c13-2中的中间偃麦草染色体同源群归属进行了分析。经过对373对SNP标记引物的筛选,结果发现只有定位于4Ee染色体上的5对引物(2对定位于4EeS,3对定位于4EeL)对Zh5-a2-1和Zh5-c13-2扩增产物的高分辨率溶解曲线与中国春明显不同,而与中5和中国春-二倍体长穗偃麦草双二倍体CS/THE相似(图4,表1)。由此说明,蓝粒代换系Zh5-a2-1和Zh5-c13-2中的中间偃麦草染色体都与二倍体长穗偃麦草的4Ee染色体同源,应为同属第四同源群的4St。由此,将蓝粒代换系Zh5-a2-1和Zh5-c13-2分别命名为SubZh5-4St(4B)和SubZh5-4St(4D)。本实验结果说明来自中5的4St染色体上带有控制蓝粒性状的基因。
2.2.4 蓝粒代换系中的中间偃麦草染色体变异
蓝粒代换系SubZh5-4St(4B)和SubZh5-4St(4D)的亲本之一是八倍体小偃麦中5,即两个代换系中的中间偃麦草染色体直接来自中5。因此,本研究对蓝粒代换系和中5中的中间偃麦草染色体进行了FISH比较分析。结果显示,在中5中含有7对中间偃麦草的染色体,但并没有与蓝粒代换系中FISH带型相同的(图5a,图2b,d)。比较发现,在中5中只有一对中间偃麦草的染色体(图5a,箭头所示的add3)在短臂近着丝点区有一个pAs1的杂交信号,长臂无信号,与代换系中的中间偃麦草染色体的信号位置及染色体形态都很相似,但在短臂末端还有一个很强的pSc119.2杂交信号。由此推测,蓝粒代换系中的中间偃麦草染色体可能是由这对染色体演变而来的。为了证明这一推测,本研究对代换系SubZh5-4St(4D)的蓝粒姊妹株进行了大量鉴定。结果发现姊妹株Zh5-c13-1的染色体数也是2n=42,GISH鉴定也显示有一对中间偃麦草的染色体,即也是一个二体代换系。但是,FISH分析却显示Zh5-c13-1中的这对中间偃麦草染色体是异形的,其中一条与中5的add3染色体的FISH结果相同,即除了在短臂近着丝点区有一pAs1的红色杂交信号外,在短臂末端也有一个很强的pSc119.2绿色杂交信号。而另一条则与其姊妹系SubZh5-4St(4D)中的FISH信号相同,即只在短臂近着丝点区有一pAs1的红色杂交信号,在短臂端部没有pSc119.2的绿色杂交信号(图5b,箭头所示)。由此推测,蓝粒代换系中的一对中间偃麦草染色体在杂交选育过程中发生了结构变异,是由中5中的一对染色体(图5a中的add3)发生短臂端部缺失后形成的。另外,在Zh5-c13-1中被代换的两条小麦染色体也不是一对,而分别是一条4B和一条4D(图5b),即是一个杂合二体代换系。

a:蓝粒代换系Zh5-a2-1染色体用Texas Red-5-dCTP标记的假鹅观草基因组(St)DNA作探针(红色)的GISH检测结果;b:与图a同一细胞用Fuorescein-12-dUTP标记的中间偃麦草基因组(EeEeSt或EeEbSt)DNA作探针(绿色)的GISH检测结果;c:蓝粒代换系Zh5-a2-1染色体用Texas Red-5-dCTP标记的二倍体长穗偃麦草基因组(Ee)DNA作探针的GISH检测结果(无杂交信号);d:与图c同一细胞用Fuorescein-12-dUTP标记的中间偃麦草基因组(EeEeSt或EeEbSt)DNA作探针(绿色)的GISH检测结果;e:蓝粒代换系Zh5-a2-1染色体用Texas Red-5-dCTP标记的百萨偃麦草基因组(Eb)DNA作探针的GISH检测结果(无杂交信号);f:与图e同一细胞用Fuorescein-12-dUTP标记的中间偃麦草基因组(EeEeSt或EeEbSt)DNA作探针(绿色)的GISH检测结果。箭头示一对中间偃麦草染 色体。
a:GISH identification of chromosomes in blue-grained substitution line Zh5-a2-1 using Texas Red-5-dCTP labelled genomic DNA ofPs.stipifolia(St) as probe (red). b. GISH identification of the same cell as subfigure a using Fuorescein-12-dUTP labelled genomic DNA ofEt.intermedia(EeEeSt or EeEbSt)as probe (green); c: GISH identification of blue-grained substitution line Zh5-a2-1 using Texas Red-5-dCTP labelled genomic DNA of diploidEt.elongata(Ee)as probe (no hybridization signal); d:GISH identification of the same cell as subfigure c using Fuorescein-12-dUTP labelled genomic DNA ofEt.intermedia(EeEeSt or EeEbSt)as probe (green);e. GISH identification of blue-grained substitution line Zh5-a2-1 using Texas Red-5-dCTP labelled genomic DNA of diploidEt.bessarabica(Eb) as probe (no hybridization signal); f. GISH identification of the same cell as subfigure e using Fuorescein-12-dUTP labelled genomic DNA ofEt.intermedia(EeEeSt or EeEbSt)as probe (green). The arrows indicate the pair ofEt.intermediachromosomes.
图3 蓝粒代换系Zh5-a2-1中中间偃麦草染色体的染色体组来源分析
Fig.3 Originated genome analysis ofEt.intermediachromosomes in blue-grained substitution line Zh5-a2-1

a:位于4EeS的SNP标记4E05的热熔解峰图;b:位于4EeS的SNP标记4E05的高分辨率熔解曲线;c:位于4EeL的SNP标记4E16的热熔解峰图;d:位于4EeL的SNP标记4E16的高分辨率熔解曲线图。CS:普通小麦中国春;CS/THE:中国春与二倍体长穗偃麦草的双二倍体。
a:The melting peaks of the SNP marker 4E05 from 4EeS; b: The melting curves of the SNP marker 4E05 from 4EeS; c:The melting peaks of the SNP marker 4E16 from 4EeL; d: The melting curves of the SNP marker 4E16 from 4EeL.CS:Common wheat Chinese Spring; CS/THE: Chinese Spring-diploidEt.elongataamphidiploid.
图4 蓝粒代换系的SNP标记鉴定结果
Fig.4 Identification of blue-grained substitution lines by SNP markers

表1 检测蓝粒代换系的特异SNP标记Table 1 Specific SNP markers for detection of blue-grained substitution lines
2.3 4St染色体特异SSR标记的筛选
为了建立更便捷的跟踪检测蓝粒代换系中中间偃麦草染色体(4St)的分子标记方法,本研究分别以中国春和中5为对照,利用450对小麦的SSR标记引物对代换系进行了鉴定。结果发现在4对引物的扩增产物中存在两个蓝粒代换系与中5一致而与中国春不同的多态性片段(图6),它们都来自第四同源群,分别定位于小麦的4A和4B染色体上(表2)。这些SSR标记可作为蓝粒代换系的特异分子标记跟踪检测4St染色体。

a:中5有丝分裂中期染色体的FISH分析,add1~add7表示7对中间偃麦草染色体,箭头示一对add3染色体;b:代换系Zh5-c13-1有丝分裂中期染色体的 FISH分析,箭头示一对异形中间偃麦草染色体(add3)。FISH探针分别为用Fuorescein-12-dUTP标记的pSc119.2(绿色)和用Texas Red-5-dCTP标记的pAs1(红色)。
a:FISH analysis of mitotic metaphase chromosomes in Zhong 5,the add1-add7 mark seven pairs of chromosomes fromEt.intermedia,the arrows indicate a pair of add3 chromosomes; b:FISH analysis of mitotic metaphase chromosomes in substitution line Zh5-c13-1,the arrows indicate a pair of heteromorphicEt.intermediachromosomes (add3). The Fuorescein-12-dUTP labelled pSc119.2 (green) and Texas Red-5-dCTP labelled pAs1 (red) were used as FISH probes.
图5 中5和蓝粒代换系Zh5-c13-1染色体的FISH比较分析
Fig.5 FISH comparative analysis of chromosomes in Zhong 5 and blue-grained substitution line Zh5-c13-1

表2 检测蓝粒代换系的特异SSR标记Table 2 Specific SSR markers for detecting blue-grained substitution lines
3 讨 论
以往的研究表明,十倍体长穗偃麦草[13]、百萨偃麦草[15]和栽培一粒小麦[17]的蓝粒基因都定位于第四同源群染色体,而本研究也证明蓝粒代换系的蓝粒性状是由中间偃麦草的4St染色体控制的。由此看来,在小麦族植物中,决定籽粒蓝色糊粉层的主效基因位点可能比较普遍地存在于第四同源群染色体上,它们很可能有共同的起源。
虽然长穗偃麦草的Ee染色体组与假鹅观草的St染色体组有很近的亲缘关系[25],但基因组水平上的差异是可想而知的。本研究利用二倍体长穗偃麦草的SNP标记成功地鉴定了蓝粒代换系中来自中间偃麦草的4St染色体。在使用的373对SNP标记引物中,有34对来自于4Ee连锁群[21],结果只有5对引物可用于鉴定4St代换系,有效率仅为14.7%。尽管有效率较低,但在St组分子标记尚缺乏的情况下,作为一种替代办法,用Ee组的SNP标记鉴定St组染色体在一定程度上还是可行的。另外,SNP标记虽然具有数量大、分布广、分辨率高等许多SSR标记无法比拟的优点,但对仪器设备和实验技术的要求都相对较高,实验成本也较高。因此,为了建立鉴定蓝粒代换系的简便方法,本研究对小麦的SSR标记进行了筛选。结果获得了4个可在小麦背景下特异性识别4St染色体的SSR标记,它们可用于蓝粒代换系的准确鉴定。
中国春缺-四体的农艺性状并不好,本研究之所以选择其作为创制代换系的亲本,主要是参考了张学勇等[27]创制小麦异源代换系的缺体回交法,期望将与缺-四体中的缺体染色体部分同源的偃麦草染色体定向地导入到小麦背景中,以提高获得部分同源染色体代换系的频率。结果获得的两份代换系材料虽然都是部分同源染色体之间的代换,即4St分别代换了4B和4D,但其中的SubZh5-4St(4B)产生于中5 × N4BT4A组合,属于定向代换,而SubZh5-4St(4D)则产生于中5×N7BT7D组合,属于非定向代换。这个结果对于可代换的21对小麦染色体来说,较随机代换具有明显的定向代换倾向。但获得的代换系材料还较少,缺-四体杂交/回交法诱导定向代换系的有效性尚需进一步验证。
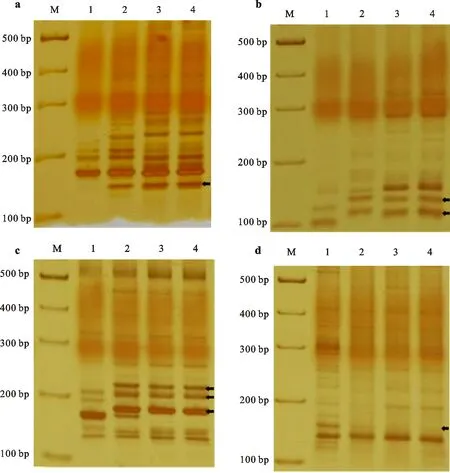
a、b、c、d分别为SSR标记gwm4、gwm251、gwm192和gwm495的扩增结果。泳道M、1、2、3和4分别为 DNA marker、中国春、中5以及蓝粒代换系SubZh5-4St(4B)和SubZh5-4St(4D)。箭头示目标条带。
a, b, c and d represent the amplification profiles of SSR markers gwm4, gwm251, gwm192 and gwm495, respectively. The lanes M, 1, 2, 3 and 4 represent DNA marker, Chinese Spring, Zhong 5, SubZh5-4St(4B) and SubZh5-4St(4D),respectively.The arrows indicate the target bands.
图6 蓝粒代换系特异SSR标记扩增图谱
Fig.6 Amplification profiles of blue-grained substitution lines with specific SSR markers
本研究获得的两份蓝粒代换系的亲本,八倍体小偃麦中5和中国春缺-四体都不是蓝色籽粒,中5的原始亲本中间偃麦草也不是蓝色籽粒。同样,产生蓝粒后代最多的长穗偃麦草也不是蓝粒,但却产生了各种类型的蓝粒后代[8,13]。已知小麦的蓝粒性状是由于在糊粉层中积累了大量的蓝色花青素,但至今对蓝色糊粉层的形成和调控机制尚属未知。在植物中,天然存在的花青素有数百种,代谢途径非常复杂。不同的基因型和不同的内外环境都可产生不同的花青素组合,从而产生不同颜色的植物组织或器官。已知蓝色小麦的糊粉层中主要有4种花青素[3],但对其相关基因的表达和调控机制还知之甚少。因此,非常有必要开展深入研究,以阐明蓝色糊粉层形成的分子机制和调控网络,为蓝粒小麦的分子设计育种和特色食品或保健品的开发提供理论依据。
