岷县生荒地和熟地育成当归苗抗逆生理特性的差异
2019-11-18白刚郭凤霞陈垣袁洪超肖婉君
白刚,郭凤霞*,陈垣,2*,袁洪超,肖婉君
(1.甘肃农业大学生命科学技术学院,农学院,甘肃省中药材规范化生产技术创新重点实验室,甘肃省药用植物栽培育种工程研究中心,甘肃省干旱生境作物学重点实验室,甘肃 兰州 730070;2.甘肃省特色药用植物资源保护与利用工程实验室,甘肃省特色药材规范化可追溯栽培工程技术研究中心,甘肃中天药业有限责任公司,甘肃 定西 748100)
当归(Angelicasinensis)为伞形科当归属草本药用植物,以干燥根入药,具有补血活血等多种功效,是我国传统大众中药材品种之一[1]。当归适宜气候冷凉,降水充沛的生态环境,并适宜排水良好,有机质含量较高的黑褐类土壤[2]。当归野生资源已趋濒危,全国第四次中药资源普查仅在甘肃省岷县和宕昌县人迹罕至的高寒阴湿灌木林发现极其稀有的野生当归资源[3]。现市售当归主要为栽培品种,地处青藏高原东部边缘地带的甘肃省岷县、漳县、卓尼县和宕昌县一带气候高寒阴湿,土层深厚肥沃,降水充沛,地势陡险,土壤不宜积水,属于我国优质当归道地产区,已有1500多年栽培史,年产量和销量均占全国当归的90%以上[4],2001年被中国特产之乡推荐暨宣传活动组织委员会命名为“中国当归之乡”。在限定使用范围和剂量内当归既是保健食品又是中药材物质的中药材品种。
当归栽培期3年,第1年种子育苗,第2年种苗成药栽培,第3年留株生产种子。当归一旦进入生殖生长其肉质根逐渐木质化。早期抽薹也称未熟抽薹,即当归在成药栽培期尚未成药而植株提早通过春化作用而抽薹开花,造成药材根木质化失去药用价值的物候特征[5]。随着中药现代化和药食同源的普及,当归需求量增大,种植面积扩大。然而,适宜当归育苗的生态区域却极其狭窄,仅局限于甘肃省岷县和漳县高海拔草原区,育成种苗质量优异,根病率和早期抽薹率均低[6]。因此,长期以来供苗基地一直沿用生荒地育苗,不占耕地,是农民传统的育苗方法。然而,随着全球气候变暖,当归早期抽薹率提高。另外,重迎茬栽培导致当归根病加重,死苗率高,严重影响栽培成效[7-11]。这些问题加重了药农对生荒地育苗的主观依赖性。生荒地育苗严重破坏了天然植被和草皮生态,极不利于保护野生物种的繁衍[12]。当归道地产区生态环境建设与传统育苗方式严重矛盾,可开垦山地几乎用尽,育苗向更高海拔荒地延伸,生态环境日趋恶化[13],大面积泥石流灾害频繁,农田生态系统功能受损,恢复机制的研究显得至关重要,探寻可替代生荒地当归育苗的熟地农茬资源是需要迫切解决的关键科技问题。尽管有人利用岷县熟土进行当归设施育苗探索,发现早播搭网育苗有一定成苗率[7],熟地黄芪(Astragalusmembranaceus)茬土与生荒土盆栽育成当归苗抗逆生化基础相当[4,14],根际微生物多样性相似[5]。宕昌县日光温室黄芪茬土也较利于当归苗的生长发育[8],但设施育苗因成本高对当归生地育苗现状的改变成效甚微。近年来,张新慧等[15]调查发现,熟地农茬口影响当归产量和质量。在作物茬口特性研究基础上[16],梁伟等[17]首次在卓尼县8种作物茬口熟地进行当归育苗研究。岷县熟地黄芪与小麦(Triticumaestivum)茬口育成当归种苗抗病性较强[6]。上述研究说明当归依赖生荒地育苗的传统方式是有望通过科学研究改变的,但熟地育苗科学问题尚不明确,缺乏与生荒地育苗的系统比较。逆境是指对作物生长不利的各种综合环境因素,抗逆植物具有发达的抗氧化系统[18-19],根病率、返青率和早期抽薹率均是反映当归苗抗逆性强弱的主要指标,豌豆(Pisumsativum)具有匀田和培肥地力的双重作用。因此,在岷县系统研究生荒地和熟地豌豆-黄芪双豆轮作茬育成当归种苗抗逆特性及其差异性具有重要意义,可为筛选替代生荒地当归育苗的熟地茬口资源提供科学依据,以改变生荒地育苗的传统格局,促进当归产业的可持续化生态发展。
1 材料与方法
1.1 试验区概况及当归种子来源
当归育苗试验设在甘肃省定西市岷县禾驮乡石家台村大石沟大湾梁,属高原型大陆性季风气候,境内平均海拔2865 m,年均气温5.5 ℃,年均日照时长2214.9 h,年均降水量800 mm,无霜期123 d,最热7月平均气温16 ℃,最冷1月平均气温-6.9 ℃,平均相对湿度68%,植被覆盖良好,属典型高寒阴湿区,高寒草甸土壤类型,为当归传统道地育苗区和栽培产区。
当归生荒地(uncultivated fields, SH )和熟地黄芪茬口(cultivated residue ofA.membranaceus, HQ)育苗试验地分别设在岷县禾驮乡石家台村大石沟大湾梁山顶和半山腰,位于同一山脉,均属温带大陆性季风气候,但海拔高度、降水量和土壤及植被类型均有差异。山顶生荒地土壤类型为高寒草甸土壤,半山腰熟地农田土壤类型为灰棕壤,均适宜当归生长发育。当归育苗前对植被观察,育苗期采用地温自动记录仪(RC-4HC Data Logger,成都华恒仪器有限公司)对生荒地和熟地试验地中心地表10 cm 处温度自动记录(表1)。
育苗试验用当归种子为2016年在生荒地留苗地3年生当归种株采集的种子,经甘肃农业大学陈垣教授鉴定为当归,挂凉保藏备用于生荒地和熟地黄芪茬当归育苗试验。

表1 2016年9月18日至2017年9月17日岷县生荒地与熟地当归育苗田地表温度比较Table 1 Comparison of annual soil surface temperatures in the uncultivated and cultivated fields for A. sinensis seedling cultivation in Min County, during September 18, 2017 to September 17, 2018
1.2 岷县熟地豌豆-黄芪茬口培育及当归育苗
在岷县禾驮乡石家台村半山腰熟地农田2013-2014年当归繁种结束,2015年种植豌豆品种陇豌2号匀田倒茬,2016年采用种子播种蒙古黄芪(A.membranaceusvar.mongholicus)育苗,培育黄芪茬口,以确保茬口土壤均匀一致。2017年当归播种育苗。黄芪茬口培育试验采用随机区组设计,3次重复,即在上年豌豆匀田基础上,2016年4月10日将试验地按坡度划分为3个区组(3次重复),小区面积4.8 m2(4.0 m×1.2 m),区组间隔0.5 m,四周设地埂,地埂宽0.5 m,每小区施用有机肥1 kg(施用量2083.3 kg·hm-2),采用开沟条播法播种蒙古黄芪种子,播深3 cm,行距10 cm,播种后耙平,以培育黄芪茬口,每小区种子播种量75 g(折合播量156.3 kg·hm-2),黄芪育苗期田间管理同大田。黄芪种苗采挖期2016年10月10日按小区用叉锨逐区采挖收获,维持小区自然状态越冬,对小区按坡度走向分别编号黄芪茬(HQ1、HQ2、HQ3),备用于次年当归熟地育苗。
熟地黄芪茬口当归育苗试验于2017年6月1日在上述已培育的茬口进行,按小区捡拾石砾及草根后,每小区施有机肥2.5 kg,用铁锨翻耕后耙平,每小区当归种子播种量为25 g(折合播量52.0 kg·hm-2),采用撒播方法播种,播种后覆盖麦草保墒增温,覆盖厚度以遮住土壤为宜。
1.3 岷县生荒地当归育苗
生荒地当归育苗试验于2017年6月1日开挖草皮后碎土捡拾草根,用铁锨翻耕后耙平,育苗方法与熟地育苗方法一致,采用随机区组设计,3次重复,即按坡度走向划分为3个区组(3次重复),除生境不同外,小区面积、每小区施用有机肥量、当归种子播种量、播种育苗方法均与熟地黄芪茬口完全一致。
上述生荒地和熟地黄芪茬口当归育苗期均不施除草剂和农药,其他农事田间管理均与大田一致。
1.4 当归种苗抗逆指标测定
当归育苗结束,2017年10月7日田间采挖种苗时对生荒地和熟地黄芪茬3个重复小区分别随机取样20苗,每处理共取样60苗,立即拌鲜土放至采样箱内低温带回实验室,备用于病株统计和抗逆生理指标测定。其余种苗拌鲜土扎把越冬贮藏,备用于次年成药栽培期返青成活率和早期抽薹率的测定。
当归种苗细胞膜稳态性按照Guo等[19]的方法以测定相对电导率和氨基酸泄漏率衡量,渗透调节能力以测定可溶性糖和可溶性蛋白质含量衡量。当归种苗脂质过氧化程度以测定丙二醛(malondialdehyde, MDA)含量和组织自动氧化速率(autoxidation rate, AR)衡量。AR测定5 h后的丙二醛含量,以每小时MDA含量的平均增加量来表示组织自动氧化速率,用于衡量活性氧自由基(reactive oxide species, ROS)对细胞的氧化攻击程度[18-19]。采用氮蓝四唑(nitro-blue tetrazolium, NBT)光化还原法测定超氧物歧化酶(superoxide dismutase, SOD)活性,采用愈创木酚显色法测定过氧化物酶(peroxidase, POD)活性,采用紫外吸收法测定过氧化氢酶(catalase, CAT)活性,测定在240 nm处每分钟光密度的变化量[18-19]。上述指标每重复小区测定3次,每处理共测定9次。
1.5 当归种苗返青成活率和早期抽薹率的测定
种苗返青结束按重复小区依次统计返青成活株数和早期抽薹株数,最后分别计算返青率和早期抽薹率。
返青成活率(regreen-survival rate)=(移栽后返青成活株数/移栽总株数)×100%
早期抽薹率(early-bolting rate)=(早期抽薹株数/返青总株数)×100%
1.6 数据统计分析
采用Excel 2003进行处理平均数间的t检验并制图。
2 结果与分析
2.1 岷县生荒地和熟地豌豆匀田后黄芪育苗田植被和生境比较
岷县禾驮乡石家台村大石沟大湾梁生荒地和熟地生境差异较大,生荒地地势陡险(坡度30°),植被覆盖度好,半山腰熟地农田以农作物为主,地势较为平缓(坡度20°)。尽管生境有一定差异,但生荒地和熟地农田四周地埂均以艾草(Artemisiaargyi)为优势种。图1显示,山顶生荒地当归育苗区属典型高寒草甸植被,以草本植物为优势种群,种群结构复杂,物种多样性丰富,与处于同一山脉半山腰的熟地农田豌豆轮作黄芪育苗地具有明显差异,熟地豌豆茬口黄芪育苗田由于定期杂草管理,植物种群结构单一,黄芪优势种生态位突出。由于生荒地大气湿度大,当归种子播种后较熟地出苗率高,群体长势良好,但熟地种苗个体生长优势较强,在2017年极端干旱条件下也获得一定数量的当归种苗(图1)。
2.2 岷县生荒地和熟地育成当归种苗根系相对电导率和氨基酸泄露率的比较
生荒地和熟地豌豆-黄芪茬育成当归种苗根系相对电导率和氨基酸泄漏率虽均具有一定差异,但差异性均未达到显著水平(P>0.05),黄芪茬育成当归种苗相对电导率更低(图2)。然而,在生荒地育成的当归种苗根系氨基酸泄露率更低。熟地黄芪茬较生荒地育成当归种苗根系相对电导率下降15.29%(t=0.926,P>0.05),但氨基酸泄露率提高9.92%(t=1.518,P>0.05)。
生荒地和熟地黄芪茬育成当归种苗根系可溶性糖含量和可溶性蛋白质含量虽有一定差异,但差异性也未达到显著水平(P>0.05),黄芪茬育成当归种苗可溶性糖含量与生荒地育成种苗的水平基本一致,但可溶性蛋白质含量低于生荒地的水平,说明黄芪茬熟地与生荒地育成当归种苗糖类渗透调节能力一致,但蛋白质渗透调节能力稍有差异。生荒地育成种苗的蛋白质渗透调节能力更强。与生荒地育成种苗相比较,熟地黄芪茬育成当归种苗可溶性糖含量仅提高0.44%(t=0.040,P>0.05),但可溶性蛋白质含量却下降23.68%(t=1.240,P>0.05)(图2)。
2.3 岷县生荒地和熟地育成当归种苗根系丙二醛含量与组织自动氧化速率的比较
熟地黄芪茬育成当归种苗根系丙二醛含量和根系组织自动氧化速率均较生荒地育成种苗的水平提高,但两者差异性不显著(P>0.05),黄芪茬与生荒地育成当归种苗MDA含量水平差异性很小,但生荒地种苗AR较低,说明熟地育成当归种苗与生荒地育成当归种苗根系膜脂质抗氧化能力略有差异,生荒地育成种苗的抗氧化能力更强,使根系MDA含量处于较低水平(图3)。与生荒地育成种苗相比较,黄芪茬熟地育成当归种苗MDA含量仅提高2.06%(t=0.478,P>0.05),而根系AR提高75.67%,但因熟地区组间变异程度大,变异系数48.14%,使两者差异性未达到显著水平(t=1.542,P>0.05)。
2.4 岷县生荒地和熟地育成当归种苗根系抗氧化酶活性的比较
不同生境条件下育苗对当归苗根系抗氧化酶活性具有一定影响(图4)。熟地黄芪茬口条件下培育的当归种苗根系SOD和POD活性均较强,CAT活性较弱,但与生荒相育成种苗间的差异性均不显著(P>0.05),熟地育成当归种苗根系SOD活性较生荒地的水平提高7.15%(t=0.909,P>0.05),POD活性提高33.96%(t=0.601,P>0.05), CAT活性却降低16.92%(t=0.903,P>0.05)。熟地育成种苗个体差异性较大,抗氧化酶类活性重复小区间的变异度均大于生荒地育成种苗的水平。
2.5 岷县生荒地和熟地育成当归种苗根病发病率和病情指数的比较
不同生境条件下育成当归种苗越冬后种苗根部发病率均在25%~35%(图5)。黄芪茬口熟地条件下培育的当归种苗根部病害发病率相对较低,较生荒相育成种苗下降10.71%(t=0.802,P>0.05),病情指数下降25.00%(t=1.375,P>0.05),但各小区种苗发病株数差异性较大,导致小区间变异度大,降低了不同来源差异显著性。

图5 生荒地和熟地育成当归种苗根病率和病情指数的比较Fig.5 Comparison of root disease rate and disease index in A. sinensis seedlings from the uncultivated and cultivated fields
2.6 岷县生荒地和熟地育成当归种苗移栽返青率和早期抽薹率动态的比较
不同土壤条件下育成当归种苗春季移栽成药栽培返青成活率和返青株早期抽薹率的变化动态差异均不显著(图6)。黄芪茬口熟地培育的当归种苗移栽后返青势强,至返青结束返青率始终高于生荒地种苗的水平,终返青率较生荒地育成种苗增加3.13%(t=0.402,P>0.05),但返青株早期抽薹率却有所提高,平均早期抽薹率较生荒地种苗的水平提高5.32%(t=0.869,P>0.05),但各小区返青株中早期抽薹率差异性大,导致小区间变异度较大,两者差异性未达到显著水平。
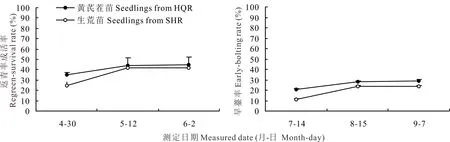
图6 生荒地和熟地育成当归种苗返青成活率和早期抽薹率的比较Fig.6 Comparison of regreen-survival rate and early-bolting rate in A. sinensis seedlings from the uncultivated and cultivated fields
种苗返青成活率与其根系CAT呈正相关,与POD活性的正相关达到显著水平(P<0.05),与SOD活性的正相关达到极显著水平(P<0.01),说明种苗抗氧化酶类活性的增强可提高当归种苗返青成活率,SOD作用更大,POD次之,CAT相对较弱。早期抽薹率与返青率呈极显著正相关(P<0.01),与抗氧化酶活性也均呈正相关,相关程度依次为POD>SOD>CAT,与POD和SOD间的正相关性达到显著水平(P<0.05)。种苗丙二醛含量与组织自动氧化速率呈极显著正相关(P<0.01),MDA含量愈高,AR越大,对种苗返青及生长发育越不利(表2)。
3 讨论
3.1 熟地豌豆-黄芪茬育苗可维持当归苗根系膜系统的稳态性
相对电导率和氨基酸泄露率是衡量植物膜系统完整性的重要指标,细胞膜稳态性差,膜系统易被氧化损伤,引起电解质泄露,相对电导率增大[18-19]。蛋白质的不稳定性可使氨基酸泄漏率增加[14]。植物在逆境下产生的大量活性氧自由基会引起膜脂质过氧化,进而损伤膜系统,MDA是膜脂过氧化的最终产物,浓度过高有毒害作用,但低浓度具有信号诱导生物体增强抗逆性的功能,MDA含量与膜稳态性和蛋白质稳定性相结合可衡量植物抗逆性,组织自动氧化速率是单位时间内MDA产生的速率,可直接反映细胞膜脂质过氧化反应的程度[18-19]。本研究中,当归种苗组织自动氧化速率与MDA含量呈极显著正相关进一步说明了两者在抗逆性评价体系中的内在联系,生荒地育成当归苗蛋白质的稳定性更强,而熟地育成当归苗膜系统稳态性更佳,因为生荒地育成苗根系氨基酸泄漏率和组织自动氧化速率略低,而熟地育成苗根系相对电导率和MDA含量略低,说明不同来源当归种苗抗逆生理机制可能存在一定差异。2017年6-10月育苗期间降水不均匀,在6-7月正值当归播种出苗期干旱少雨,但育苗后期降水多,可能导致熟地当归苗逆境压力势更大,但种苗并没有受到严重伤害,因为根系通过消耗可溶性蛋白质提高抗逆性,MDA处于较低浓度范围,有利于激活其抗氧化系统机制,才得以维持种苗生物膜系统的稳态性。

表2 生荒地和熟地育成当归种苗抗逆生化指标与返青率和早期抽薹率的相关性分析Table 2 Relation of physicochemical stress resistant indicators to the regreen rate and early-bolting rate of A. sinensis seedlings from the uncultivated and cultivated fields
注: X1, 相对电导率。** 表示相关性达极显著水平(P<0.01), * 表示相关性达显著水平(P<0.05),n=18,R16,0.05=0.468,R16,0.01=0.590。
Note: X1, relative conductivity. ** means great significant correlation atP<0.01, * means significant correlation atP<0.05,n=18,R16,0.05=0.468,R16,0.01=0.590.
3.2 熟地豌豆-黄芪茬育苗可改善当归苗根系糖类渗透调节能力
可溶性糖担负着渗透调节作用,在植物旺盛光合作用期间通过渗透调节作用维护着细胞渗透势、细胞水分平衡和光合作用[19]。一般植物的抗逆性与可溶性糖的大小呈正相关[20]。极端逆境影响植物的生长发育,可溶性蛋白作为保护性物质也参与植物细胞的渗透调节作用,可溶性蛋白含量与植物的抗逆性呈正相关[21-22]。本研究表明,豌豆-黄芪双豆间迎茬条件下当归种苗可溶性糖和可溶性蛋白含量均接近生荒地种苗的水平,但可溶性糖较生荒地种苗提高0.11%,可溶性蛋白质含量下降4.58%,尽管差异性未达到显著水平,但可说明在遇干旱气候条件下,生荒地育苗对当归种苗有更适宜的环境优势,环境压力较小,也可说明熟地农茬口可造就环境压力势,双豆茬轮作造就的环境压力势在一定程度上提高了当归种苗的渗透调节功能及生理调控保护作用,从而有利于增强当归种苗的抗逆性。
3.3 熟地豌豆-黄芪茬育苗可增强当归种苗根系SOD和POD酶促抗氧化系统并降低根病率
生物自由基损伤理论证明当植物受到各种非生物因子胁迫时,细胞内活性氧(ROS)包括超氧阴离子(O2·-)、过氧化氢、羟基自由基(·OH)和单线态氧(1O2)等积累过多,其高度的氧化活性可破坏细胞膜结构,使植物抗逆性下降,导致植株发病甚至死亡,抗逆植物体内可进化出高效抗氧化机制,及时调控体内ROS以减轻这种氧化伤害[18]。SOD主要将最具有毒性的O2·-分解为H2O2和H2O,H2O2进一步由POD和CAT协同清除[19]。抗氧化系统发达的植株可抵抗氧化胁迫,有效保护膜系统[21-22]。本研究中,熟地豌豆-黄芪茬与生荒地育成当归种苗根系内MDA含量基本一致,但抗氧化酶类活性存在一定差异。熟地育成种苗SOD和POD活性均高于生荒地种苗的水平,CAT活性却低于生荒地的水平,尽管差异性未达到显著水平,但也可以反映出熟地黄芪茬育苗环境对当归具有一定环境胁迫势,因为CAT是抗逆性最弱的抗氧化酶类,逆境条件下一般CAT最先受到影响,易失活。尽管黄芪茬育成当归种苗CAT下降了16.92%,但也维持了一定的CAT活性,较生荒地种苗下降程度未达到显著水平,这与金彦博等[23]在岷县蚕豆(Viciafaba)-黄芪茬口基础上研究的结果相似,进一步说明黄芪茬口培育的当归种苗可以增强SOD和POD维持根系的生物抗氧化功能。这从种苗返青成活率得到进一步解释,即豌豆-黄芪茬口育成当归种苗较生荒地育成种苗早期抽薹率虽增加5.32%,但返青成活率提高3.13%,这可能是由于熟地育苗期间种苗受到干旱胁迫,加速了种苗的自我繁殖保护能力的根源所在,因为早期抽薹也是植物应对环境变迁的抗逆表现,营养生长期的干旱胁迫可诱发当归物候的转变,即从营养生长及早进入生殖生长,完成生活史。抗氧化系统发达的种苗返青成活率和早期抽薹率均高的结果也证明了这一点。由于土壤是植物生长生存的环境,植物可以改变土壤的生态环境,进而影响后茬植物的生长发育。植物对茬口的适应也是一种抗逆表现[24]。一般在田间最大持水量的60%范围内,土壤含水量越高,土壤微生物的活性愈强,从而利于植物的生长发育[25]。虽然当归重迎茬在土壤中积累了自毒物质,豆科作物豌豆和黄芪均属于绿肥作物,可能对降解当归自毒物质、促进种苗生长发育均具有重要作用。
4 结论
茬口形成的因素较为复杂,但茬口特性的好坏最终表现在后茬作物的产出性能上,尽管熟地豌豆-黄芪间茬口育成当归种苗早期抽薹率较生荒地苗提高5.32%,但根病率下降10.71%,病情指数下降25.00%,由于根系SOD和POD活性较强,返青率提高7.52%,成药率提高11.39%,有效补偿了早期抽薹率偏高造成的产出损失,说明熟地适宜作物轮作茬口育成当归种苗具有与生荒地种苗相似但有差异的抗逆生理机制,豌豆-黄芪倒茬轮作可替代生荒地进行当归育苗。一般6月中旬麦草覆盖育苗,生长期100 d即可采挖种苗,拌土入窖越冬,次年春季移栽大田进行成药栽培,但熟地育苗期需加强水肥管理,底肥强化有机肥,根据天气情况及时遮阳为宜。
致谢:岷县禾驮乡郎银忠和李雪芳提供试验地和合作,陇西县巩昌镇郭志军、郭天明和郭太平协助试验地田间管理。中国科学院大学博士生陈翔,兰州大学硕士生郭爱峰,甘肃农业大学硕士生梁伟、金彦博、许美玲、王雪琴参与育苗试验,硕士生郭一青、王小琴、陈永中、刘兰兰,本科生甄世伟参与指标测定。在此一并致谢。
