Rabdosinate调节生长素极性运输蛋白PIN1、PIN3和PIN4抑制拟南芥幼苗根生长
2019-11-15郇兆蔚
李 芃 郇兆蔚 丁 兰
(西北师范大学生命科学学院,兰州 730070)
次生代谢产物是地球上含量最丰富的有机化合物的宝库,约有20余万种,萜类是其中的最庞大的一群,包括单萜、倍半萜、二萜、三萜、四萜及多萜[1]。迄今,从动物、植物和微生物中已分离鉴定出4万余种萜类化合物[2]。萜类在植物中广泛存在,并在植物生长发育过程中承担着重要功能。植物的代谢产物可分为初生代谢产物和次生代谢产物。初生代谢的萜类在保证生物膜的完整性,生物膜系统上的电子传递,光合作用以及作为植物激素调节植物生长发育等方面发挥着重要作用[3]。次生代谢的萜类大量存在于植物的根、茎、叶中,在生物农药、食品添加剂及药物开发利用方面显示了重要的商业价值,并已有广泛研究报道[4~5]。研究显示,次生代谢的萜类还是第二大类化感物质,具有重要的化学生态作用。植物体内含量巨大及种类繁多的萜类通过自然挥发、根系分泌或枯枝落叶的淋溶降解等方式释放于自然环境之中,成为生态环境演变的重要参与者,并对其它植物的生长发育产生重要影响,发挥植物化感作用,参与植物种间竞争[6]。植物化感作用除了对自然生态产生重要影响,还在农业生产(作物轮作等)及外来植物入侵方面显示其重要的研究价值[7~9]。尽管化感作用的研究历史很长,但由于所涉及的可变因子多,其过程复杂及研究手段的局限性,导致相关研究还比较滞后,目前对大多数次生代谢产物的化感作用机制还很不明确[3]。近年来,模式植物拟南芥的各种突变体及转基因株系为化感作用分子机制研究提供了极好的研究平台,取得了较大进展。
植物次生代谢产物对生境中其它植物的生长发育产生显著影响,产生植物化感作用。主要表现在3个方面:生长促进,生长抑制以及向重力性干扰。目前的文献报道主要集中于生长抑制作用[10~13]。研究显示,不同的萜类化合物显示了不同的生长抑制机制。例如,细胞膜渗漏,线粒体解偶联,线粒体氧吸收抑制,细胞周期阻滞,有丝分裂指数降低,细胞DNA损伤,种子储藏蛋白活性抑制等[8,10~11,14]。近年来,研究者更加关注次生代谢产物对植物体内植物激素的调控干预作用,更多集中于化合物对生长素途径的影响[12~13]。众所周知,不同植物激素的信号途径相互关联并形成复杂的信号网络,对植物生长发育发挥着决定性的调控作用,而生长素是其中的核心调控因子,主导植物生长发育的全过程[15]。因此,着眼于次生代谢产物对植物激素尤其是生长素途径的干预作用,对于化感作用机制的阐释有重要意义。
香茶菜属植物中含有极为丰富的对映—贝壳杉烷型二萜,报道有500余种[16]。大量的该类二萜可随枯枝落叶进入自然生境,产生化感效应。之前,我们实验室对不同分子骨架类型的该类二萜的化感潜能进行了评估,发现leukamenin E和weisiensin B比rabdosin B及epinodosin有更高的植物毒性[17~20]。而rabdosin B和epinodosin则显示了双重效应,即,低浓度具有生长促进作用,高浓度抑制生长。发现低浓度rabdosin B和epinodosin可促进莴苣种苗根尖分生区细胞的有丝分裂以及根尖细胞的伸长生长,高浓度条件下可诱导根尖细胞周期阻滞并显著降低根尖细胞的有丝分裂指数以及抑制细胞伸长生长[10~11],这提示可能与植物激素调节相关联。进一步检测发现,leukamenin E处理后的拟南芥幼苗根部生长素含量显著升高,证实该化合物介导了植物根组织的生长素调节[18]。为了阐述该类二萜介导生长素的调节途径,揭示其植物化感作用的分子机制,本文报道另一种具有螺环内酯型分子骨架的对映—贝壳杉烷二萜rabdosinate调节生长素极性运输转运子PIN1、PIN3和PIN4抑制拟南芥幼苗根生长发育的规律。
1 材料与方法
1.1 材料
对映—贝壳杉烷二萜rabdosinate(1α,6,11β,15β-tetraacetoy-6,7-seco-7,20-olide-ent-kaur-16-en)由本实验室从甘肃产蓝萼香茶菜(Isodonjaponicavar.galaucocalyx)中分离得到,并采用波谱学方法(红外、质谱及核磁共振等)对其结构进行了确定[21]。实验用rabdosinate单体用溶剂DMSO配制成100 mmol·L-1母液,DMSO培养基中终浓度不高于0.1%,当DMSO低于该浓度时对拟南芥幼苗生长无显著性影响,并对实验用转基因株系中所检测的相关蛋白量及其定位无显著影响(数据未显示)。
野生型拟南芥(Col-0)购于ABRC(ArabidopsisBiological Resource Center);突变体pin1、pin3和pin4以及转基因株系DR5::DR5:GFP、PIN1::PIN1:GFP、PIN3::PIN3:GFP、PIN4::PIN4:GFP由中国科学院遗传与发育研究所李传友研究员惠赠。上述拟南芥突变体及转基因株系均为Col-0生态型背景。
1.2 方法
拟南芥种子经2% NaClO表面消毒5 min, 用无菌水漂洗5次后点播于1/2 MS培养基上,4℃条件下春化3 d,然后转移至培养箱内培养3 d,选择长势一致的拟南芥种苗置于含有不同浓度(40、60、80 μmol·L-1)rabdosinate的培养基上继续培养。野生型及突变体幼苗于5 d后取出拍照、固定并测定拟南芥幼苗主根长以及统计侧根数和侧根原基数。根据Zhang的分类办法[22],侧根原基(包括侧根)分为4个时期:A期≤3层细胞;B期>3层细胞并未突破主根表皮,C期突破主根表皮并其长度<0.5 mm,D期突破主根表皮并且其长度≥0.5 mm。
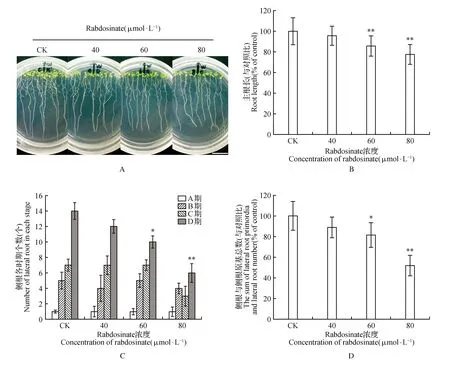
图1 Rabdosinate对拟南芥根生长的影响(标尺=1 cm) A.图示rabdosinate对拟南芥幼苗生长的影响;B.rabdosinate抑制拟南芥主根生长;C.rabdosinate对拟南芥侧根各时期(A~D期)影响;D.rabdosinate减少拟南芥侧根原基和侧根之和数 *P<0.05;**P<0.01 下同。Fig.1 Rabdosinate effects the growth of Arabidopsis roots(Scale bars=1 cm) A.Effects of rabdosinate on the growth of Arabidopsis seedlings; B.Rabdosinate inhibits Arabidopsis root growth; C.Effects of rabdosinate on the number of lateral roots in A-D stage in Arabidopsis seedlings; D.Rabdosinate reduces the total number of lateral root primordia and lateral root *P<0.05; **P<0.01,the same as below.
转基因株系DR5:DR5:GFP、PIN1::PIN1:GFP、PIN3::PIN3:GFP、PIN4::PIN4:GFP于药物处理24和48 h后取出,在Leica荧光显微镜(Leica,DM 4000B)下拍照,每处理组至少15苗。
实验所用基本培养基为MS培养基。拟南芥幼苗均培养于1/2 MS添加1%蔗糖、0.56%琼脂,pH值5.8的培养基上。培养温度22±2℃,光周期为16 h光照/8 h黑暗。
1.3 数据统计分析
各组数据均用均值±标准差来表示,数据使用SPSS软件进行单因素方差分析(ANOVA),各组数据之间运用LSD方法进行差异显著性检验,P<0.05为差异显著,P<0.01为差异极显著。使用Image J软件(https:imagej.nih.gov/ij/)对获得的荧光图片进行荧光强度测定。
2 结果与分析
2.1 Rabdosinate对野生型拟南芥幼苗根生长的影响
本实验首先采用40、60和80 μmol·L-1的rabdosinate处理3天苗龄的野生型拟南芥幼苗,培养5 d后检测rabdosinate对幼苗生长发育的影响,发现rabdosinate对幼苗根生长显示了一定的选择性抑制效应(图1),而对胚轴生长没有显著影响(数据未显示)。从图1可以看出,低浓度处理组(40 μmol·L-1)对幼苗根主根及侧根生长基本无影响,其主根长(图1:A~B)、侧根和侧根原基数(图1:D)与对照相比均无显著性差异。而较高浓度处理组(60和80 μmol·L-1)中rabdosinate显著抑制拟南芥幼苗主根长(图1:A~B),减少侧根和侧根原基数(图1:D)。Rabdosinate在60和80 μmol·L-1浓度时,对主根抑制率均达到极显著水平(P<0.01)对侧根和侧根原基之总和数的抑制率则分别为显著水平(P<0.05)和极显著水平(P<0.01)。从图1C可以看出,rabdosinate主要对C期和D期的侧根原基有明显的抑制作用,而对A期和B期侧根原基无明显的影响,表明对侧根起始有延迟作用。
2.2 Rabdosinate对拟南芥幼苗根尖部生长素分布的影响
DR5启动子是监测植物Auxin响应的非常有用的手段。研究证实,生长素信号在一定浓度范围内(高于DR5响应阈值或低于DR5响应饱和值),DR5启动子可反映组织或细胞的生长素水平,具有正相关关系[23~25]。
为了探讨rabdosinate抑制主根生长及侧根发育是否与根尖生长素分布的改变相关联,我们进一步通过转基因报告株系DR5::DR5:GFP检测了rabdosinate对根尖部生长素分布的影响。将rabdosinate处理DR5::DR5:GFP报告株系幼苗24和48 h后,将其根尖置于荧光显微镜下观测报告基因荧光强度及分布情况,组织中绿色荧光强度的变化可表征生长素含量的变化。观察及测试结果如图2。从图2可以看出,报告基因的绿色荧光蛋白分布于根尖部分生区,位于小柱细胞和静止中心及其周围细胞。48 h对照组(5 d苗龄)幼苗根分生区荧光强度明显高于24 h对照组(4 d苗龄)的荧光强度(图2A:a,e),表明生长素在该区域的含量与其苗龄相关。处理组与对照组相比,发现rabdosinate可明显升高幼苗根尖部荧光强度,并随着处理浓度升高荧光增强(图2),此外,48 h的高浓度处理组幼苗根尖部绿色荧光分布范围比其对照组有扩展趋势(图2A:e~h)。这些结果表明,rabdosinate对拟南芥主根及侧根的抑制效应可能与其改变根部生长素含量及分布有关。
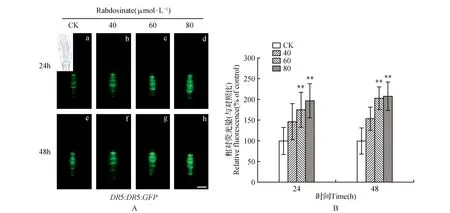
图2 Rabdosinate对拟南芥根部DR5-GFP表达的影响(标尺=50 μm) A.不同处理组的拟南芥根尖DR5-GFP表达的代表性图像;B.不同处理组的拟南芥根尖DR5-GFP的相对荧光强度Fig.2 Effects of rabdosinate on the expression of DR5-GFP in Arabidopsis roots(Scale bars=50 μm) A. Representative images of the expression of DR5-GFP in rabdosinate-treated roots; B. Relative fluorescence intensity of DR5-GFP in rabdosinate-treated roots
2.3 Rabdosinate对拟南芥pin1、pin3、pin4突变体幼苗根生长发育的影响
生长素在植物生长发育中发挥着极为关键的调控作用,生长素的一个重要特征是它的极性运输,极性运输不仅对生长素进行转运,更是实现生长素在植物组织中的梯度分布的必需途径,在此过程中,不同类型的生长素转运蛋白对于生长素在植物组织中的梯度分布起着关键作用。生长素PIN蛋白家族是一类生长素极性运输外输载体蛋白,有8个成员,PIN1、PIN2、PIN3、PIN4和PIN7属一个亚族,定位于细胞膜,将细胞内的生长素转运至膜外,担负组织中生长素浓度梯度形成与维持的功能[26]。
为了求证根尖部生长素分布变化是否是与rabdosinate对生长素极性运输PIN蛋白的调节作用相关,我们进一步用rabdosinate(40~80 μmol·L-1)处理3种生长素外输载体的突变体pin1、pin3和pin4,6 d后检测突变体主根长以及侧根和侧根原基数量,并与野生型拟南芥相比较(图3)。40~80 μmol·L-1的rabdosinate处理突变体后,仅80 μmol·L-1处理组与其未处理组相比有抑制轻微效应,但未显示显著性差异。与野生型相比,rabdosinate处理的突变体(pin1、pin3和pin4)的主根未显示显著的抑制效应,表明pin1、pin3和pin4突变后导致rabdosinate对主根抑制的恢复作用(图3:A)。Rabdosinate对突变体侧根数的影响表现为:40 μmol·L-1的rabdosinate对突变体pin1、pin3和pin4侧根及侧根原基之和数没有显著影响,这与野生型的响应一致,而60和80 μmol·L-1的rabdosinate对突变体侧根及侧根原基之和数有减少效应,但与野生型相比,80 μmol·L-1处理组的减少幅度明显降低(图3:B)。上述实验结果表明rabdosinate参与了生长素极性运输转运子PIN1、PIN3和PIN4的调节作用。
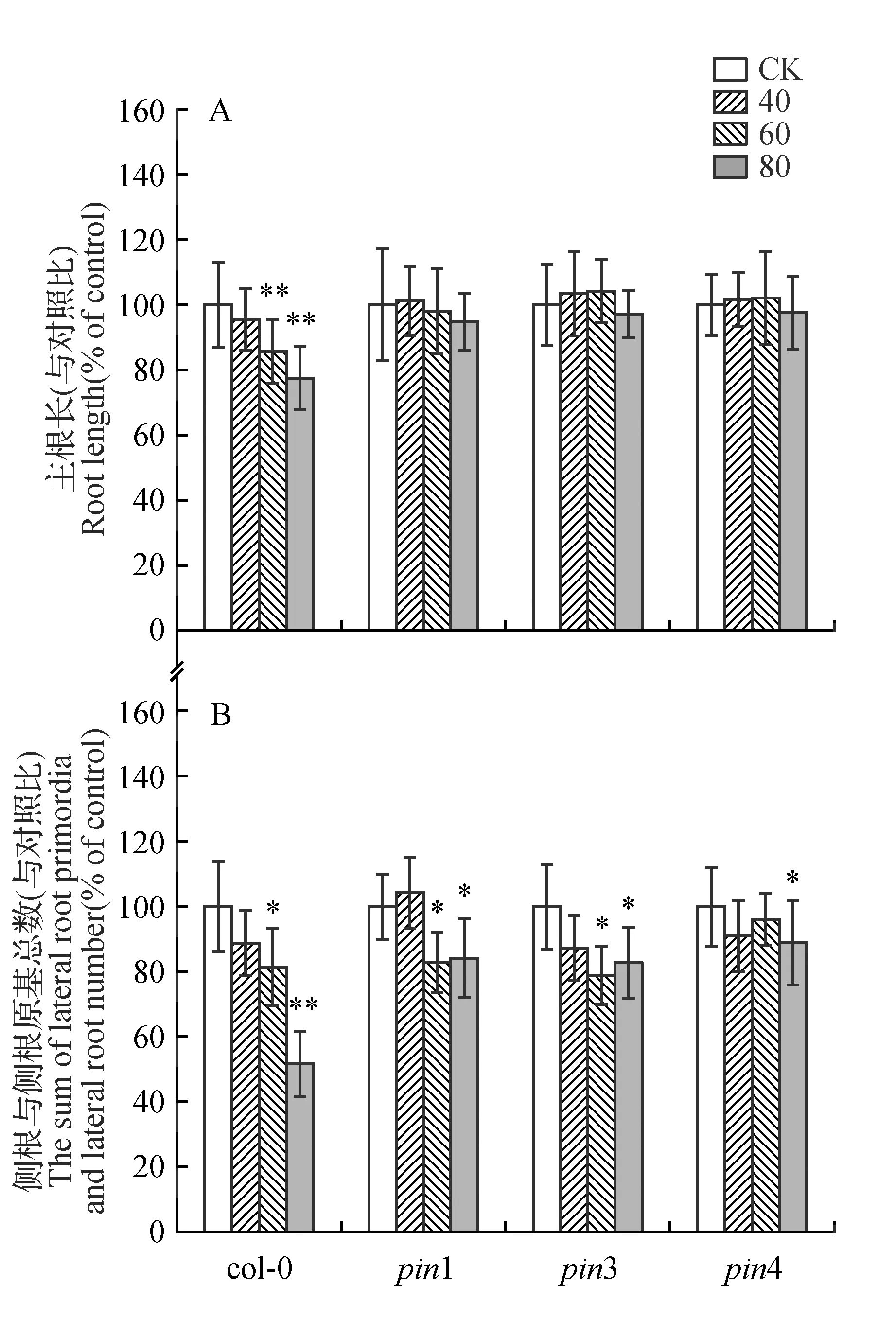
图3 Rabdosinate对拟南芥pin1、pin3和pin4突变体根生长的影响 A.Rabdosinate影响野生型和突变体拟南芥主根生长的比较;B.Rabdosinate影响野生型和突变体拟南芥侧根及侧根原基总数的比较Fig.3 Effects of rabdosinate on the root growth of Arabidopsis mutant pin1,pin3 and pin4 A.Comparison of the root growth between wild type and mutant seedlings after treatment with rabdosinate; B.Comparison of the total number of lateral root primordia and lateral roots between wild type and mutant seedlings after treatment with rabdosinate
2.4 Rabdosinate对拟南芥幼苗根部生长素极性运输转运子PIN1、PIN3和PIN4表达的影响
为了了解rabdosinate如何对生长素极性运输转运子PIN1、PIN3和PIN4产生影响,本研究通过荧光显微镜观察转基因株系PIN1::PIN1:GFP、PIN3::PIN3:GFP和PIN4::PIN4:GFP幼苗根部融合蛋白PIN1-GFP、PIN3-GFP和PIN4-GFP的荧光分布,以表征生长素极性运输外输载体蛋白PIN1、PIN3和PIN4在根尖部细胞的定位情况。PIN1主要定位于幼苗根部中柱和内皮层细胞;PIN3定位于中柱鞘、中柱及小柱细胞,而PIN4定位于静止中心及其下侧细胞。上述3种转基因株系幼苗在含有不同浓度的rabdosinate培养基上培养24和48 h后,发现其根部中柱细胞的融合蛋白PIN1:GFP的荧光强度明显增强(图4A:a~d,a1~d1和B),表明PIN1在中柱细胞分布丰度增加;但rabdosinate减弱融合蛋白PIN3-GFP在小柱细胞(图4A:e~h,e1~h1和C)以及PIN4:GFP在静止中心及其下部细胞的荧光强度(图4A:i~l,i1~l1和D),表明可抑制PIN3和PIN4在这些区域的定位丰度,并显示了时间依赖性和浓度依赖性。
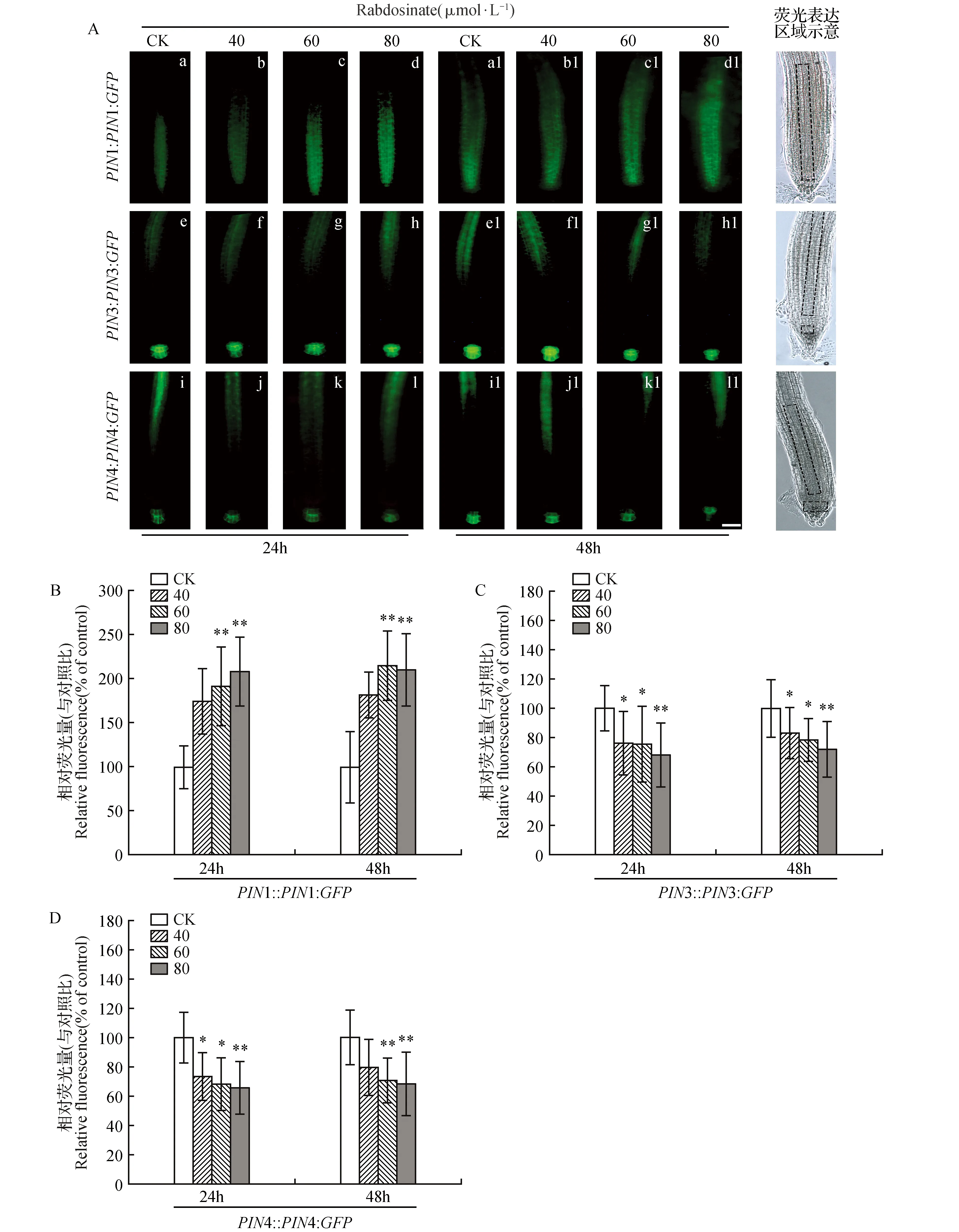
图4 Rabdosinate对拟南芥转基因株系根尖部融合蛋白PIN1-GFP、PIN3-GFP和PIN4-GFP丰度的影响(标尺=50 μm) A.Rabdosinate影响PIN1、PIN3和PIN4丰度的代表性图像;B~D.Rabdosinate处理后根尖融合蛋白的相对荧光强度Fig.4 Effects of rabdosinate on the abundance of PIN1-GFP,PIN3-GFP and PIN4-GFP in the root tip of Arabidopsis transgenic lines(Scale bars=50 μm) A. Representative images of fusion protein PIN1-GFP,PIN3-GFP and PIN4-GFP in the root tip of Arabidopsis transgenic lines after treatment with rabdosinate; B-D. Relative fluorescence intensity of fusion protein in the root tip of Arabidopsis transgenic lines after treatment with rabdosinate
3 讨论
文献报道显示,大多受试植物次生代谢产物对其它植物生长发育的影响表现为高低不同的植物毒性(生长抑制),某些化合物在低浓度时对植物生长有促进效应[27~28]。然而,目前对植物毒性的作用机制的阐释还不明确,其分子机制的研究还很有限。近年来,植物激素调节植物生长发育的机制取得了重大进展,研究证实了生长素在植物生长发育的核心作用,参与调节了极为广泛的发育过程,明确了生长素极性运输所形成的组织内生长素浓度梯度在参与调控植物器官发生、发育和向性反应等生理过程中的关键作用[15,29]。既然植物次生代谢产物对植物生长发育的影响主要表现为生长抑制或促进效应,那么,它们是否介导了生长素途径,是否干预生长素极性运输相关载体而影响生长素在组织中的浓度或分布,进而对植物生长产生影响。本研究提出了对映—贝壳杉烷二萜rabdosinate参与生长素极性运输调节影响拟南芥根生长的实验证据。
之前,我们对甘肃产不同种类香茶菜属植物进行了植物化学研究,得到了多种对映—贝壳杉烷二萜[21,30]。进一步研究显示,几种不同分子构型的受试二萜对测试植物的生长抑制或促进作用均显示与分生区细胞的有丝分裂及细胞伸长相关联[10~11,17]。其中的二萜leukamenin E不仅抑制拟南芥主根生长同时也减少侧根数量[18],还升高幼苗根部生长素含量[18]。研究证实,生长素促进侧根原基细胞分化并调控侧根形态建成,而生长素梯度信号则是诱发侧根原基起始的主要因素[31],同时生长素也具有维持根分生区细胞的有丝分裂活性及其之后的快速伸长分的功能[15]。在本研究中,通过观察分析拟南芥报告株系DR5::DR5:GFP的融合蛋白丰度,证实rabdosinate也可升高根尖分生区生长素量,显示了生长素在根尖部区域的累积效应(图2:A),与leukamenin E对株系DR5::DR5:GFP幼苗根的处理结果相类似(图片未显示)。尽管rabdosinate与leukamenin E属不同的分子骨架类型,rabdosinate为螺环内酯型,leukamenin E为C-20未氧化型,但上述结果表明它们对拟南芥生长抑制作用可能具有相似的调节机制。
为了解释rabdosinate引起根尖生长素累积现象,明确是否与生长素极性运输相关,我们进一步检测rabdosinate对株系PIN1::PIN1:GFP、PIN3::PIN3:GFP和PIN4::PIN4:GFP幼苗根部融合蛋白荧光定位及强度。结果显示,rabdosinate可增强融合蛋白PIN1-GFP在幼苗根尖中柱细胞的荧光强度,但rabdosinate降低融合蛋白PIN3-GFP在小柱细胞和PIN4-GFP在静止中心及其下部细胞的荧光强度,表明生长素极性运输外输载体蛋白PIN1的定位丰度增加,而PIN3和PIN4的定位丰度减少(图4:A)。PIN1主要具有参与生长素在根部的向顶运输的功能[32]。本实验中,PIN1丰度的增加可增强生长素向根尖部运输,导致尖部细胞生长素量增多;PIN3具有参与小柱细胞区域生长素横向运输的功能[33],PIN3丰度的减少会导致生长素向小柱细胞两侧的分配减弱而小柱细胞生长素积累。PIN4具有维持静止中心及其下方细胞生长素浓度并保持根部生长素浓度梯度的作用。最直接的证据是突变体pin4根尖分生区生长素水平升高,并且其生长素浓度梯度被显著改变[34]。由此推测,本文中PIN4分布的减少不仅干扰根尖分生区生长素浓度梯度的维持,并可能对生长素在根尖部的积累也有贡献。根分生区生长素浓度及浓度梯度的维持是该部位细胞正常有丝分裂、伸长生长及侧根产生所必需。研究显示,在拟南芥幼苗发育过程中,高浓度外源生长素的加入会导致幼苗主根生长抑制,而低浓度外源生长素则促进主根生长,同时也影响侧根发生,直观地表明了生长素及其浓度梯度的改变对根生长发育产生的干扰。显然,rabdosinate对生长素极性运输外输载体蛋白PIN1定位丰度的增加以及对PIN3和PIN4丰度的减少极可能是导致了根尖部生长素积累的关键因素,而根尖部生长素的积累不仅影响细胞分裂及细胞伸长,也会破坏根尖生长素浓度梯度,干扰侧根的发生及生长,最终导致了拟南芥幼苗根生长的抑制效应。另外,突变体实验进一步证实了rabdosinate介导了对PIN1、PIN2和PIN3的调节作用:与对照组相比,rabdosinate对3种突变体pin1、pin3和pin4主根生长的抑制作用消失,而对侧根数量的减少有明显的恢复效应。
综上,本研究表明对映-贝壳杉烷二萜rabdosinate可能通过影响生长素外输载体PIN1、PIN3和PIN4的定位丰度,升高生长素在根分生区的浓度,打破根部生长素浓度梯度平衡,导致主根生长抑制及侧根数量减少。本研究仅对3种PIN蛋白进行了检测。目前已发现多类生长素运输载体,例如,外输载体PIN家族成员(PIN1~PIN8)及内输载体AUX/LAX等,至于rabdosinate是否还参与了对其它种类的生长素运输载体的调节作用,还需进一步的深入研究。此外,考虑到根尖分生区细胞具有生长素合成能力,因此,也不能排除rabdosinate可促进生长素的合成,上述结论尚需进一步验证。
