青海高原披碱草属种间天然杂种的细胞学鉴定
2019-11-15路兴旺刘瑞娟窦全文
路兴旺 刘 博 刘瑞娟 窦全文*
(1.中国科学院西北高原生物研究所,中国科学院高原生物适应与进化重点实验室,西宁 810008; 2.中国科学院大学,北京 100049; 3.中国科学院西北高原生物研究所,青海省作物分子育种重点实验室,西宁 810008)
披碱草属(ElymusL.)是禾本科(Poaceae)小麦族(Triticeae)中含有物种种类最多、分布范围最广的一个属,其中包含大约150个由小麦族不同近缘属物种通过自然杂交形成的物种[1]。研究表明,披碱草属物种基因组组合由St、H、W、Y和P 5个基本基因组组成[2~3],其中St基因组来源于拟鹅观草属(Pseudoroegneria(Neveski)Löve)、H基因组来自于大麦属(HordeumL.)、W基因组来自于澳冰草属(Australopyrum(Neveski)Löve)、P组来自于冰草属(AgropyronJ.Gaertn.)、Y基因组的来源迄今尚不明确[3~4]。披碱草属物种为典型的异源多倍体物种,所有物种由上述不同的基因组组合组成。北美学者Löve依据这些物种间共享基因组以及种间可杂交的特性,将上述所有物种归并于一个大属,即披碱草属[1];然而,我国分类学家颜济依据披碱草属形态学和基因组组成研究结果将其划分为以下几个属:披碱草属(ElymusL.,基因组组合为StH)、鹅观草属(RoegneriaC.Koch,基因组组合为StY)、以礼草属(KengiliaYen et J.L.Yang,基因组组合为StYP)、弯穗草属(Campeiostachys,基因组组合为StHY)以及花鳞草属(Anthosachne,基因组组合为StWY)[5]。
披碱草属物种呈现复杂的网状进化特征,种间杂交基因渗透普遍存在[6]。Mason-Gamer等[7]发现在北美和欧亚大陆间的四倍体披碱草属间存在一定的种间杂交基因渗透现象;Wu等[8]揭示在同域分布的披碱草属物种E.alaskanus(Scribn. and Merr.) Love,E.caninus,E.fibrosus和E.mutabilis之间发生了种间杂交基因渐渗,存在双向或单向的基因流。甚至,在亲缘关系很远的物种间,披碱草属也可发生种间杂交基因渗透,如在E.repens中检测到了来自Taeniatherum属的GBSSI序列[9],以及来自Bromus和Panicum属的ITS序列[10],这在一定程度上揭示了同域分布的披碱草属种间杂交基因渗透不仅发生在近缘种间,而且在亲缘关系很远的物种间也可能发生。
青海高原是披碱草属物种多样性分布地区,该地区几乎分布所有基因组组合类型的披碱草属物种(除StWY外),该属中一些物种类型在青海高原地区具有高度变异和广泛的适应特性。该地区披碱草属不同种间杂交、甚至披碱草属物种与其他近缘属物种之间杂交基因渗透,有可能在披碱草属物种多样性以及物种遗传多样性形成中发挥着重要作用。
不同物种间通过天然杂交形成F1代杂种,是种间发生基因渗透的关键初始步骤。在本研究中,我们在对披碱草属物种野外天然杂交形成的不育材料的观察、采集基础上,利用细胞遗传学手段对杂种的基因组来源进行了鉴定分析,进一步确定了天然杂种的身份。研究结果可以为研究披碱草属物种间杂交基因渗透提供重要的依据,同时鉴定出的部分天然杂种,通过进一步的开发有可能作为生态草种或优良牧草品种,具有潜在的种质材料价值。
1 材料和方法
1.1 材料
实验材料垂穗披碱草(ElymusnutansGriseb.)和鹅观草属(RoegneriaC.Koch)杂种,垂穗披碱草和达乌力披碱草(E.dahuricusTurcz. ex Griseb)杂种以及垂穗披碱草和大颖草种(KengiliagrandiglumisKeng)杂种采自青海海晏(平均海拔3 200 m);垂穗披碱草和糙毛以礼草(KengiliahirsutaKeng)杂种,老芒麦(ElymussibiricusL.)和达乌力披碱草种间杂种以及糙毛以礼草和赖草(Leymussecalinus(Georgi) Tzvel.)杂种采自青海贵南(平均海拔3 100 m)。
1.2 方法
1.2.1 种间杂种根尖获取
由于所有正常生长的披碱草属物种种子在秋季表现落粒特性,结实正常的小穗在秋季均从主穗轴脱落,但是不育小穗依然可以保留在主穗轴上,因此在秋冬季,在野外观察披碱草穗部小穗的保留特征可以对天然杂种进行初步筛选鉴定,并统计结实率以及与同域分布披碱草物种进行形态比较,可以初步确定在形态上呈现出较多中间性状且结实率为零的个体为种间杂种。将鉴定出的个体进行整体活体采挖,移植至中科院西北高原生物所院内进行盆栽,待恢复生长一段时间后,获取新鲜生长出的次生根,供进一步细胞学鉴定利用。
1.2.2 染色体制片
取根尖后放于1.5 mL EP管中,打开管口并保持管内湿润,在8个标准大气压下的N2O气室中处理2 h,处理结束后向每管中加入500 μL卡诺固定液(无水酒精∶冰醋酸=3∶1,体积比),4℃固定30 min以上,然后将固定好的根尖置于干净的载玻片上,滴加一滴45%的醋酸,盖上盖玻片并用镊子轻轻敲打以使根尖细胞分散,接着将片子在酒精灯上微热几秒,用大拇指用力垂直压片,最后利用相差显微镜(Olympus Bx43)先在10倍镜下观察,然后选择分裂相多、染色体分散和形态清晰的片子转移至40倍镜下确定染色体形态和数目,最终,选择分裂相较好的制片于-80℃冰箱中冰冻30 min以上。
1.2.3 探针制备
重复序列由寡核苷酸序列pAs1代表[11],寡核苷酸片段pAs1和(AAG)10由生工生物工程(上海)有限公司合成,并在5′端经TAMRA(红色)或FITC(绿色)荧光基团修饰,作为荧光原位杂交(FISH)探针。
提取糙缘拟鹅观草(Pseudoroegneriastipifolia(Czern. ex Nevski) A.Löve,2n=14,StSt基因组)、布顿大麦(HordeumbogdaniiWilensky.,2n=14,HH基因组)、扁穗冰草(AgropyroncristatumJ.Gartern.,2n=14,PP基因组)以及赖草(Leymussecalinus(Georgi) Tzvel.,2n=28,NsNsXmXm基因组)基因组DNA,经120℃高温消解2 min,然后利用随机引物标记法将消解后的基因组DNA用Tetramethyl-rhodamine-5-dUTP(红色)或fluorescein-12-dUTP(绿色)标记,作为基因组原位杂交(GISH)探针。
1.2.4 荧光原位杂交
制备好的染色体制片在-80℃揭片,自然干燥;配制杂交液(50%甲酰胺、50%硫酸葡聚糖、1 mg鲑鱼精DNA、1 μL 20×SSC和10 ng探针DNA);将干燥的染色体制片放入0.2 mol·L-1溶于70%酒精的NaOH溶液中变性10 min;随后取出立即置于-20℃预冷的无水乙醇中,脱水干燥;FISH寡核苷酸探针无需变性,可以直接加入杂交液中杂交,GISH探针与杂交液混合后在沸水中热变性10分钟;接着将染色体制片放置在潮湿的密封盒中,37℃避光过夜杂交(二次杂交不需要再变性);用蒸馏水冲洗后,洗耳球吹干制片的正面,向制片中心加入10 μL含DAPI(4,6-diamidino-2-phenylindole)的抗荧光衰减封片剂封片,然后在荧光显微镜(Olympus Bx63)下观察拍照,获取的图片在Adobe Photoshop 6.0中进行调节。
2 结果与分析
2.1 垂穗披碱草和鹅观草属物种间杂种的鉴定
此杂种在垂穗披碱草居群中发现,根尖染色体计数为35条,为五倍体杂种,初步推断可能是六倍体垂穗披碱草和一四倍体披碱草物种间的杂交F1代。进一步利用顺序FISH/GISH进行细胞学分析,FISH和GISH杂交特征显示7条染色体为H基因组染色体,14条染色体为St基因组染色体,其他14条染色体为Y基因组染色体(图1:a1~a2),此杂种染色体组成为StStYYH。因为垂穗披碱草染色体组成为StStHHYY[12],所以杂种另一亲本染色体组成必然为StStYY,并且该地区同域分布有大量StStYY四倍体鹅观草属物种[13],另外杂种中St、Y基因组中pAs1和AAG杂交FISH特征与我们在前期研究中揭示的该地区垂穗披碱草和部分四倍体鹅观草染色体FISH特征具有高度相似性[12~13],由此我们判定此杂种为垂穗披碱草和一鹅观草属物种间的种间杂种。
2.2 垂穗披碱草和达乌力披碱草种间杂种的鉴定
此杂种同样发现于垂穗披碱草居群中,染色体计数为42条。利用顺序FISH/GISH鉴定结果表明,此杂种个体具有14条完整的St组染色体,13条完整的Y基因组染色体,13条完整的H基因组染色体,以及2条H基因组和Y基因组之间易位染色体(图1:b1~b3),2条易位染色体呈单倍性状态,初步判定此杂种为两个染色体组成为StStHHYY披碱草物种间的同倍体杂种。由于该地区同域分布有大量的达乌力披碱草物种,其染色体组为StStYYHH[14],达乌力披碱草物种染色体组中稳定存在一对H/Y易位染色体[14~15],杂种中鉴定出的2条单倍性状态的易位染色体的FISH和GISH特征与达乌力披碱草物种中存在的物种特异性易位染色体呈现高度一致性[14~15],另外杂种穗形介于达乌力披碱草和垂穗披碱草穗形之间。由此,我们判定此杂种为垂穗披碱草和达乌力披碱草间的种间杂种。
2.3 老芒麦和达乌力披碱草种间杂种的鉴定
此杂种根据形态学特征在老芒麦居群中发现,染色体计数为35条。由于老芒麦为异源四倍体物种(2n=28,染色体组为StStHH),推测此杂种为老芒麦和一六倍物种间的杂种,进一步利用顺序FISH/GISH进行鉴定,结果揭示此杂种具有14条完整的St组染色体,6条完整的Y组染色体,13条完整的H组染色体,以及2条H基因组和Y基因组之间易位染色体(图1:c1~c2),杂种染色体组成为StStHHY。初步推断,此杂种为四倍体老芒麦和六倍体StStHHYY披碱草物种间的杂种。同样,此杂种中2条处于单倍性状态的H和Y易位染色体与达乌力披碱草物种特异性易位染色体高度一致,由此我们判定此杂种为老芒麦和达乌力披碱草间种间杂种。
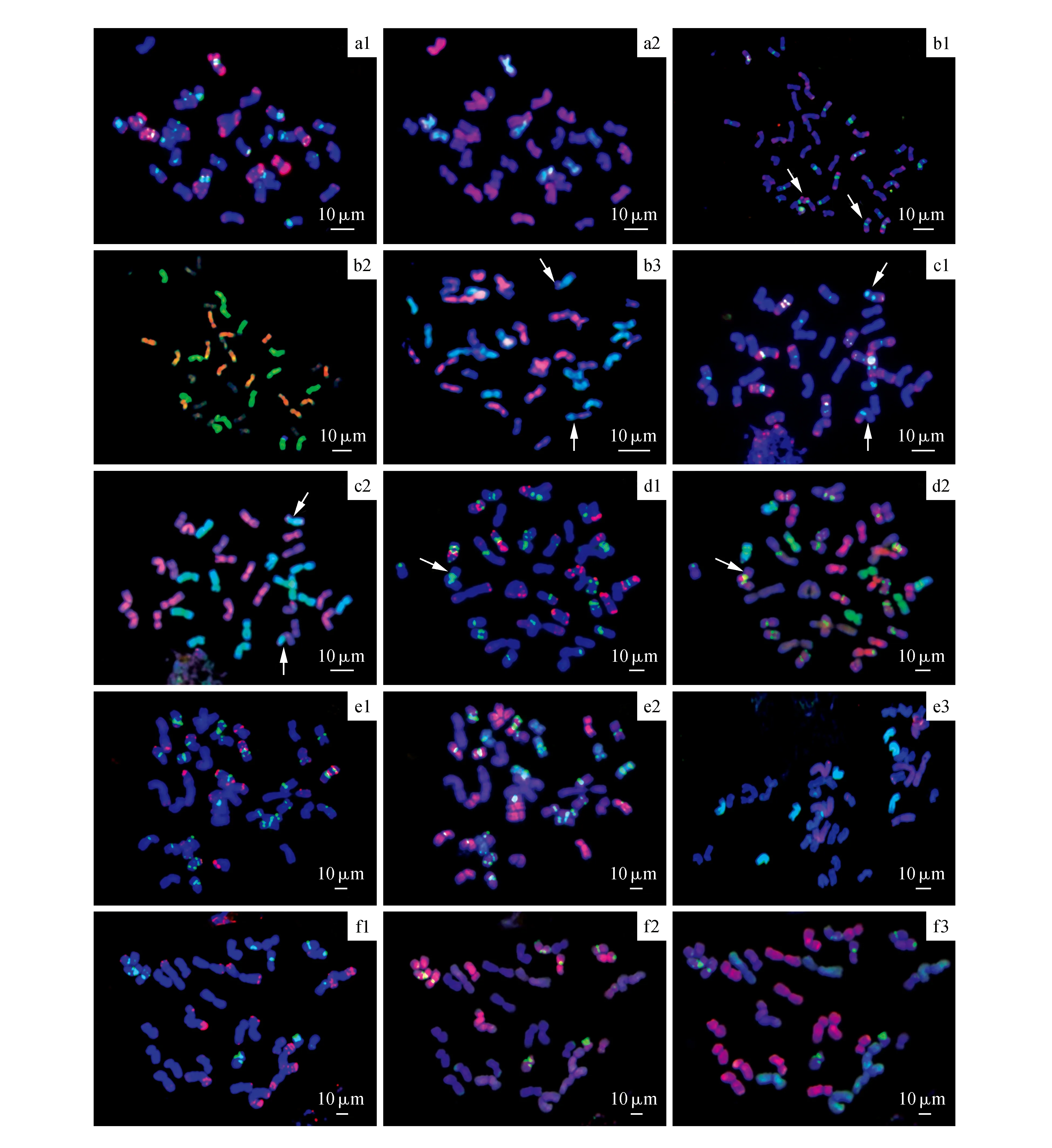
图1 披碱草属物种不同种间杂种根尖有丝分裂中期染色体的FISH和GISH鉴定 a.垂穗披碱草和鹅观草间杂种(染色体数为35);b.垂穗披碱草和达乌力披碱草间杂种(染色体数为42);c.老芒麦和达乌力披碱草间杂种(染色体数为35);d.垂穗披碱草和糙毛以礼草间杂种(染色体数为42);e.垂穗披碱草和大颖草间杂种(染色体数为42);f.糙毛以礼草和赖草间杂种(染色体数为35)a1、b1、c1、d1、e1、f1利用pAs1(红色)和(AAG)10(绿色)为探针组合进行FISH杂交;a2、b2、c2、d2、e2、f2为利用St基因组(红色)和H基因组(绿色)为探针组合的顺序GISH杂交;e3和f3为分别利用P基因组(红色)和H基因组(绿色)为探针组合、以及赖草基因组(红色)和P基因组(绿色)为探针组合的GISH杂交。箭头所示为基因组间易位染色体。Fig.1 FISH and GISH patterns on root tips mitotic metaphase cells of interspecific hybrids in Elymus a,b,c,d,e,f are hybrids between E.nutans and Roegneria,E.nutans and E.dahuricus,E.sibiricus and E.dahuricus,E.nutans and K.hirsuta,E.nutans and R.grandiglumis and K.hirsuta and L.secalinus respectively a1,b1,c1,d1,e1 and f1 are FISH patterns hybridized by probes of pAs1(red) and(AAG)10(green); a2,b2,c2,d2,e2 and f2 are sequential GISH patterns hybridized by probed St(red) and H(green) genomic DNAs; e3 and f3 are GISH patterns probed by combination of probed P(red) and H(green) genomic DNAs,and combination of probed L.secalinus(red) and P(green) genomic DNAs,respectively.The arrows indicate the inter-genomic translocations.
2.4 垂穗披碱草和糙毛以礼草种间杂种的鉴定
此杂种在垂穗披碱草和糙毛以礼草分布的中间地带通过形态学观察得到,染色体观察计数为42条,进一步利用细胞学鉴定揭示此杂种含有13条完整的St组染色体,14条完整的Y组染色体,一条St和Y易位染色体,7条H组染色体以及其他7条形态较长的染色体(图1:d1~d2),根据FISH杂交特征推断7条形态较长染色体应该为P基因组染色体,判定杂种染色体组成为StStYYHP,推测为垂穗披碱草和一染色体组成为StStYYPP物种间的杂种。由于同域伴生有大量糙毛以礼草物种,染色体组成为StStYYPP,其染色体分子核型特征与此杂种中部分St、Y和全部P组染色体FISH杂交信号呈现高度相似性[16]。由此,我们判定此杂种为垂穗披碱草和糙毛以礼草种间杂种。垂穗披碱草核型中,染色体组间易位经常发生[14,17],此杂种的一条易位染色体可能来自亲本垂穗披碱草。
2.5 垂穗披碱草和大颖草种间杂种的鉴定
此杂种通过形态学观察在大颖草居群边缘地带发现,染色体观察计数为42条。利用FISH和GISH细胞学鉴定此杂种分别具有14条St组和Y组染色体,7条H组染色体,7条P组染色体(图1:e1~e3),染色体组成为StStYYHP,推测其为六倍体StStYYHH和StStYYPP物种间天然杂种。由于在此大颖草居群周边伴生有大量垂穗披碱草物种,结合此杂种与大颖草和垂穗披碱草染色体分子核型特征[14,17]分析,我们判定此杂种为垂穗披碱草和大颖草种间杂种。
2.6 糙毛以礼草和赖草种间杂种的鉴定
此杂种在赖草居群分布生境中得到,染色体根尖计数为35条。相邻区域分布有大量的垂穗披碱草和糙毛以礼草物种,初步推测为四倍体赖草和六倍体垂穗披碱草或糙毛以礼草杂种,利用pAs1和AAG探针杂交显示此杂种染色体上AAG杂交信号较含StYH基因组的杂种显著减少(图1:f1),进一步利用H基因组和St基因组探针杂交显示无明显H基因组信号检出,7条染色体为St组染色体,显示St信号较弱的7条为Y组染色体,7条较大染色体上有微弱H基因组信号分布,其余14条染色体无任何杂交信号(图1:f2)。为进一步确定染色体的身份,利用赖草基因组和冰草基因组探针进行第三次杂交,结果表明此杂种中含有14条完整的赖草染色体和7条P基因组染色体(图1:f3)。由此,我们推定此杂种为赖草和糙毛以礼草种间杂种,染色体组成为StYPNsXm。
3 讨论
本研究揭示了3个五倍体杂种,其中StStHHY和StStYYH杂种为披碱草属内不同倍性物种间非对称杂交后代,两种杂种染色体组合类型在青海高原不同地区被发现和鉴定,StStHHY杂种被推定为四倍体老芒麦和其他未知六倍体披碱草间杂种[18~19],StStYYH杂种被推定为垂穗披碱草和短颖鹅观草(R.breviglumisKeng et S.L.)间杂种[20]。在本研究中揭示的上述两个五倍体杂种类型有可能与其他研究者报道的部分五倍体杂种来自相同的亲本物种。本文揭示的糙毛以礼草和赖草之间的五倍体杂种为一未曾报道过的类型。文中揭示的3个六倍体型杂种,其中垂穗披碱草与大颖草间杂种与相吉山[20]所报道的来自相同的亲本物种,达乌力披碱草与垂穗披碱草种间杂种有可能与相吉山[20]未能明确鉴定的一种垂穗披碱草和六倍体杂种来源相同,糙毛以礼草与垂穗披碱草间杂种未见报道。另外,结合本研究披碱草属物种间天然杂种的鉴定以及其他研究结果,我们认为在青海高原地区披碱草属物种间天然杂交高频率发生,深入系统地对该地区天然杂种进行鉴定分析,有可能发现更多披碱草物种间、乃至属间的杂种类型。
相吉山[20]在对种间杂种鉴定时,发现不同基因组间染色体有较高频率的易位发生,但是不能确定其易位来源,其认为在杂种中存在的相互易位染色体有可能是杂种形成过程中产生的,而其他易位染色体应该源自亲本物种。在本研究中杂种也有少量易位检出,而且检出的易位染色体呈单倍性状态,在杂种类型没有检测到相互易位染色体存在,对比分析其亲本物种的染色体特征,我们认为杂种中检测到的易位来自于亲本物种,因为杂种形成过程本身不能产生易位。
种间杂交基因渗透有可能是披碱草属物种和遗传多样性形成的一个重要驱动因子[21]。以礼草属多个物种是青海高原地区特有物种,而且某些物种种下有变种发生[22],本研究以及前人研究[19~20]揭示以礼草属物种大颖草及糙毛以礼草与同域分布的垂穗披碱草、无芒披碱草、乃至赖草属物种间极易产生杂交,这些结果提示种间杂交有可能在该地区以礼草属物种多样性形成中起着重要作用。垂穗披碱草(E.nutans)是高寒区最为引人关注的一个草种,且在生境上具有普遍的适应特性,在海拔1 000~5 000 m的河滩、山坡、灌丛以及高寒草地等均有广泛分布,从形态学、生化、分子、核型等不同水平上均呈现很高的变异特性[12,23~25]。垂穗披碱草自交结实率高,为自花授粉植物[26],但是遗传多样性检测揭示居群内多样性高于居群间多样性[23~25]。在本研究中揭示垂穗披碱草可以与多个不同的披碱草属物种发生种间杂交,这种结果提示种间杂交有可能是高原地区垂穗披碱草物种遗传多样性形成的一个重要驱动因素,局部地域分布的垂穗披碱草多样性形成有可能受到同域分布披碱草物种杂交基因流的强烈影响。种间杂交形成杂种是种间基因渗透的初始步骤,进一步回交形成后代才是种间基因渗透的关键环节。在本研究中我们只是得到了有关披碱草属不同种间可以形成杂种的证据,而进一步利用实验手段揭示回交的程度及在多样性形成中的作用是非常必要的。
披碱草属中众多物种为青海高原天然草场优质牧草,尤其是老芒麦和垂穗披碱草在高寒地区表现出抗逆性强、生物量高、适口性强等优点,被驯化为优良牧草品种在饲草生产中广泛应用。本研究揭示的老芒麦、垂穗披碱草和其他物种间的杂种,有可能作为特殊种质材料进行进一步育种利用。利用非对称杂交杂种,如StStHHY或StStYYH杂种,进一步与四倍亲本物种回交,在后代中有可能选育出携带有不同染色体的单体附加材料,可以作为优异基因定位、发掘的良好材料。本研究鉴定出的糙毛以礼草和赖草间杂种具有St、P、Y、Hs和Xm各一套染色体组,对这个材料直接加倍,有可能获得结实性良好、遗传相对稳定的材料。糙毛以礼草在沙丘与草地过渡生态环境中具有很好适应性,且种子结实率高;而赖草抗旱性强、根茎系统强大,但是结实率极低。因此,对其杂种类型的进一步开发利用,有可能培育出抗逆性强大、结实率较高的可用于高寒地区生态恢复利用的新型草种。
