不同叶龄杉木人工林叶碳氮化学计量及其稳定同位素的海拔梯度变化特点
2019-11-04王亮曹小青孙孟瑶李培玺滕臻徐小牛
王亮,曹小青,孙孟瑶,李培玺,滕臻,徐小牛
安徽农业大学林学与园林学院,安徽 合肥 230036
叶是植物生理上最活跃的植物器官(Gupta et al.,2012),其与环境接触面积最大,特性会随着特定环境做出适应性变化(Schwoertzing et al.,2016);叶指挥植物中某些对生态系统有严重影响的生理活动,表现出高度的耐受性和对气候变化的不同反应(Guerin et al.,2012)。因此,长期以来与叶相关的研究备受生理生态学者的关注。碳氮作为自然界中很重要的生命元素,往往会产生分馏效应,导致碳氮同位素在不同物质之间存在差异,这为同位素技术在生态系统碳氮循环过程中的应用提供了理论基础(Natelhoffer,1988)。碳同位素 δ13C已经被开发为测量水分利用效率(WUE)的工具,因为在δ13C和WUE之间发现有强相关性(Farquhar et al.,1989)。植物氮同位素δ15N是氮循环转化的综合结果,可以间接反映陆地生态系统氮循环的特征;δ15N已被用作生态系统氮饱和状态的指示指标(Högberg,1997;Xu et al.,2010)。
海拔差异会导致生态因子,诸如温度、降水和土壤类型的变化,使生长的植物更加复杂,也导致植物碳氮同位素对海拔梯度的响应更复杂。关于碳氮同位素随海拔的变化,目前大部分研究是植物叶片δ13C随海拔梯度的变化,植物13C的分辨在一定程度上反映未来气候环境变化对植物的影响(史作民等,2004),多数研究显示C3植物的δ13C随着海拔梯度的增加而增加(Körner et al.,1988;Hultine et al.,2000;Shi et al.,2006),也有研究表明 C3植物δ13C随着海拔梯度的增加而降低(Lajtha et al.,1993)。Wang et al.(2010)研究发现,δ13C不受海拔梯度的影响。通常,δ13C主要受到植物自身(生理生态、遗传变异、基因、叶片结构)以及外部环境中的气候因子的影响(沈芳芳等,2017),所以海拔差异导致的气候因子以及植物叶片结构、生理生态的变化很大程度上会影响植物叶片δ13C含量。一般而言,植物中的氮同位素δ15N反映了广泛的生物地球化学过程,主要与氮源、吸收、同化和分配有关(Szpak,2014)。关于δ15N与海拔梯度的关系研究变化趋势也有不少,有研究表明,C3植物δ15N和海拔之间具有一定的负相关关系,可能与在冷湿环境下土壤呼吸速率降低引起土壤中轻同位素14N富集有关(Aranibar et al.,2010;Linn et al.,1984)。Amundson et al.(2003)在总结已发表的文献后认为,在全球范围内,在海拔梯度上陆地植物δ15N与气候因子具有很大的关系(与温度正相关,与降水负相关),但在局部地区,由于微环境的影响,植物δ15N随海拔的变化也可能出现正相关或不相关的情况。目前关于植物碳氮同位素对海拔响应的研究中,大部分忽视了叶龄对碳氮同位素影响,可能对研究结果有一定影响。为此,本研究在探讨植物碳氮同位素对海拔响应的基础上,把不同叶龄的叶片分开,以探讨叶龄与海拔的交互作用,使研究结果更具有准确性。
杉木(Cunninghamia lanceolata)是南方广泛种植的优良针叶用材树种,栽培面积达9.0×106hm2,在产业用材生产中发挥着重要作用(Tang et al.,2016)。本研究基于海拔梯度固定试验样地,通过分析不同海拔梯度下杉木人工林不同叶龄鲜叶碳氮磷化学计量关系以及碳氮同位素变化特征,为杉木人工林科学经营提供依据。
1 研究地概况
研究地位于安徽金寨县马鬃岭林场(31°10′N,115°40′E),总面积约 3500 hm2,森林蓄积量约1.4×105m3,森林覆被率达90%。属于北亚热带温润性季风气候,年均降水量约1315 mm,年均温度约16.6 ℃,海拔350-1500 m,坡度范围是25°-40°,土壤以偏酸性的黄棕壤、山地黄壤为主。森林类型多,名木古树丰富,属国家保护的树种有领春木(Euptelea pleiosperma)、香果树(Emmenopterys henryi)、连香树(Cercidiphyllum japonicum)、大别山五针松(Pinus dabeshanensis)、野生山核桃(Juglans cathayensis)、三尖杉(Cephalotaxus fortunei)、银雀树(Tapiscia sinensis)、黄山木兰(Magonlia cylindrica)等。此外,20世纪70年代,该地区营造了大量杉木人工林,目前保存面积超过1500 hm2。
选取 4个不同海拔的杉木人工林作为研究对象,分别为750、850、1000、1150 m。每个海拔设置3个20 m×20 m的样地,共12个样地。各海拔杉木人工林的林分生长状况如表1所示。
2 研究方法
2.1 杉木鲜叶的采集与处理
2018年7月底对每块样地进行每木检尺,测量树高和胸径,根据平均标准木法选取 4-5棵标准木,用高枝剪选取杉木下部朝南方向、新鲜、没有变黄的叶片,按照不同叶龄(当年生、1年生、2年生和3年生)进行采集,把同一块样地中同叶龄的叶片混合装袋并带回实验室处理,共采集 48个样品。带回实验室后将样品放入信封中,置于65 ℃的烘箱中烘干至恒重,然后用自动球磨仪磨碎后过100目筛,待测。
利用同位素质谱仪(Thermo Scientific Delta V Advantage,Germany)测定稳定碳氮同位素的丰度值(δ13C和δ15N,‰),计算公式如下:
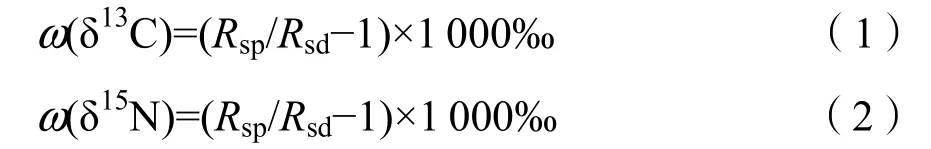
式中,R为13C/12C或者15N/14N;Rsp为叶片样品的R值;Rsd为标准物质的R值;碳同位素以PDB(Pee Dee Belemnite)为标准,氮同位素以标准大气中的N2为标准。
利用元素分析仪(EA3000,Italy)来测定样品碳(C)和氮(N)含量。采用湿式消化法获得待测液,用自动流动注射分析仪(FIA Star 5000 Analyzer,FOSS,Sweden)测定样品磷(P)含量。
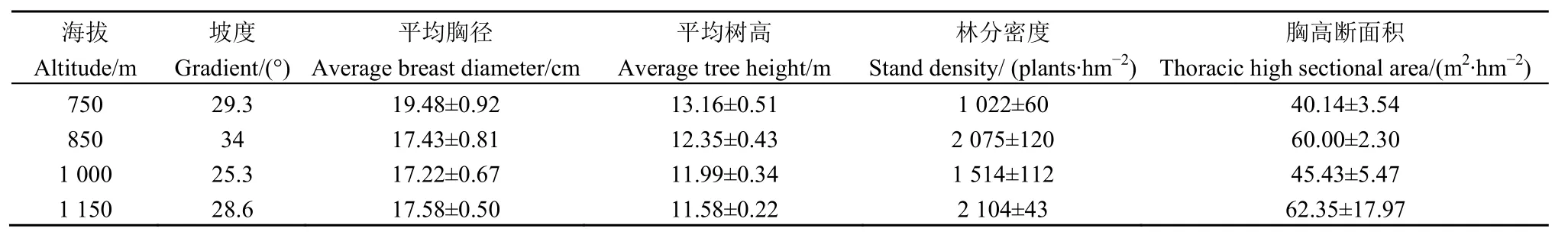
表1 杉木试验林生长状况Table 1 Growth status of the experimental plantations of Cunninghamia lanceolata
2.2 数据处理
运用Excel 2013和R 3.5.1软件对数据进行统计分析,用Origin Pro 2018软件制图,图表中的数据为平均值±标准误。采用方差分析和LSD法分析海拔和叶龄对各因子的影响,采用Pearson相关系数来检验各因子之间的相关性,采用回归分析建立各因子之间的回归模型。
3 结果与分析
3.1 不同海拔、叶龄对δ13C和δ15N的影响
如表2所示,海拔对杉木叶片δ13C含量的影响极显著(P<0.01)。当年生叶中,海拔1150 m叶片δ13C含量(-29.40‰)显著高于750 m的δ13C含量(-30.49‰),且随着海拔梯度的增加叶片δ13C含量逐渐增加;海拔对其他各叶龄叶片影响不显著(图1A)。叶龄对叶片δ13C含量影响不显著(P=0.388),在每个海拔梯度下,各叶龄叶片δ13C含量变化并无显著规律,总体而言,随着叶龄的增加,叶片δ13C含量先降低再增加,表现为当年生(-30.06‰)>3年生(-30.36‰)>1年生(-30.56‰)>2年生(-30.59‰)(图 1A)。海拔和叶龄的交互作用对叶片δ13C含量影响不显著(P=0.945)。

表2 海拔和叶龄及其交互作用对杉木叶碳氮同位素影响的方差分析Table 2 Results of variance analysis of altitude and foliar age and their interactions on carbon and nitrogen isotopes in foliage of Cunninghamia lanceolata
海拔对叶片δ15N含量影响不显著(P=0.092),同一叶龄叶片在不同海拔梯度下其 δ15N含量变化无显著规律,总体上,随着海拔的增加δ15N含量先降低后增加,表现为 1150 m(-4.58‰)>750 m(-5.07‰)>850 m(-5.16‰)>1000 m(-5.23‰)(图1B)。叶龄对δ15N含量影响显著(P<0.05),同一海拔不同叶龄叶片之间 δ15N含量均随着叶龄的增加而降低,其中海拔850 m当年生叶片δ15N含量(-4.73‰)显著高于 3年生叶片 δ15N含量(-5.60‰),1150 m当年生叶片δ15N含量(-4.23‰)显著高于2年生和3年生叶片δ15N含量(-4.81‰,-4.82‰),750 m和1000 m各叶龄叶片δ15N含量无显著差异。总体而言,随着叶龄的增加叶片δ15N含量逐渐降低,且当年生叶片δ15N含量(-30.06‰)显著高于3年生叶片δ15N含量(-30.36‰)(图1B)。海拔和叶龄的交互作用对叶片 δ15N含量无显著影响(P=0.77)。
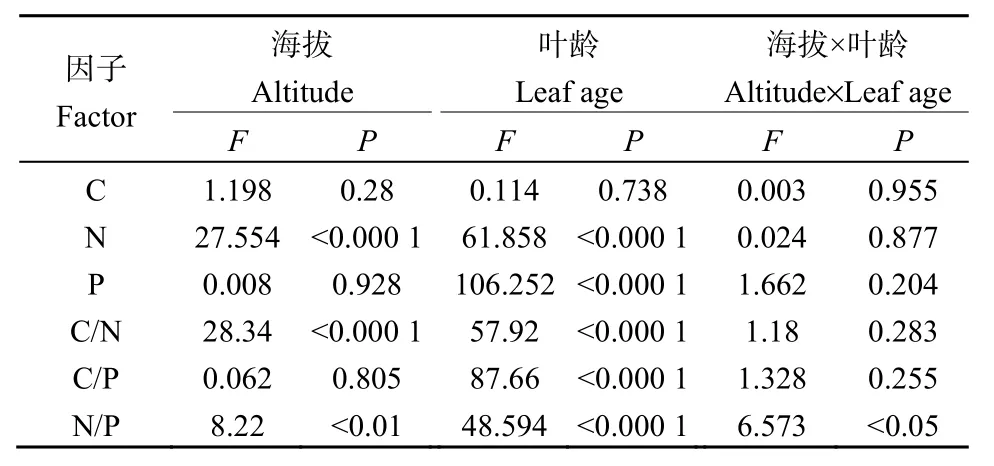
表3 海拔和叶龄及其交互作用对杉木叶片碳氮磷含量及其化学计量比影响的方差分析Table 3 Results of the variance analysis of altitude and foliar age and their interaction on concentrations of C, N, P and their stoichiometric ratios in foliage of Cunninghamia lanceolata
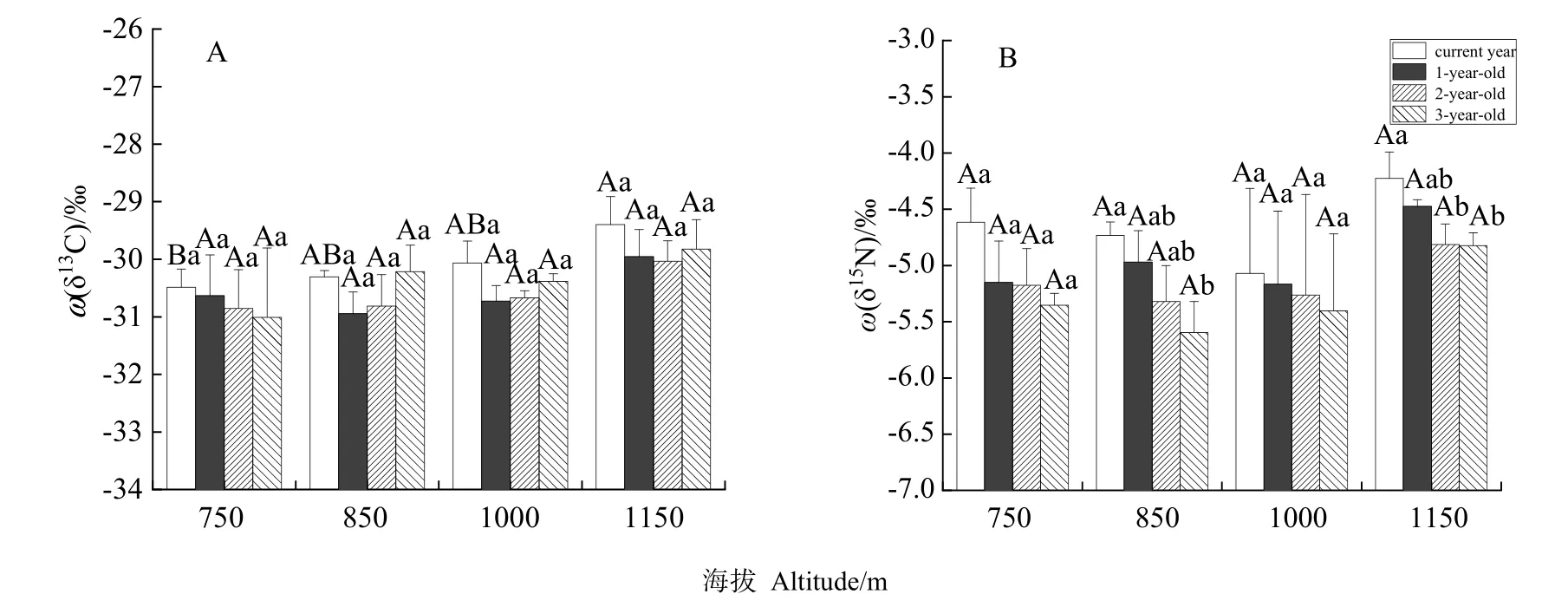
图1 不同海拔和叶龄杉木叶片δ13C、δ15N之间的差异Fig. 1 Differences of δ13C and δ15N in foliage of Cunninghamia lanceolata between different altitudes and foliar ages
3.2 不同海拔、叶龄对C、N、P含量及其化学计量比的影响
如表3所示,海拔和叶龄及其交互作用对叶片C含量均无显著影响(P=0.28,P=0.738,P=0.955)。海拔和叶龄对叶片 N含量影响均极显著(P<0.0001,P<0.0001),当年生叶片 N 含量在海拔变化上未产生显著差异,其他叶龄叶片在海拔变化上产生显著差异(表4)。总体上,海拔750 m的叶片N含量极显著高于其他海拔梯度的N含量,表现为 750 m(15.49 g·kg-1)>850 m(13.90 g·kg-1)>1000 m(13.40 g·kg-1)>1150 m(12.89 g·kg-1);同一海拔不同叶龄叶片N含量均有显著差异,呈现出随着叶龄的增加N含量逐渐降低;海拔和叶龄的交互作用对叶片 N含量无显著影响(P=0.877)。海拔对叶片 P含量无显著影响(P=0.928),叶龄对叶片 P含量影响极显著(P<0.0001),在同一海拔水平上,不同叶龄叶片差异均达到显著水平,随着叶龄的增加叶片P含量逐渐降低(表4),海拔和叶龄的交互作用对叶片P含量无显著影响(P=0.204)。
海拔和叶龄对C/N影响均极显著(P<0.0001,P<0.0001),当年生叶片C/N在海拔变化上未出现显著差异,1-3年生叶片C/N均受到海拔的显著影响,表现为随着海拔的增加叶片C/N逐渐增加(表4);同一海拔不同叶龄之间C/N均有显著差异,呈现随着叶龄的增加C/N逐渐增加;海拔和叶龄的交互作用对C/N无显著影响(P=0.283)。海拔对叶片C/P无显著影响,叶龄对叶片 C/P影响极显著(P=0.805,P<0.0001),同一海拔不同叶龄叶片C/P差异显著,随着叶龄的增加C/P逐渐增加(表4),海拔和叶龄的交互作用对 C/P无显著影响(P=0.255)。海拔和叶龄及其交互作用对 N/P均有显著影响(P<0.01,P<0.0001,P<0.05),且 N/P在海拔上的变化趋势与C/P一致,海拔降至1000 m时开始上升(表4),
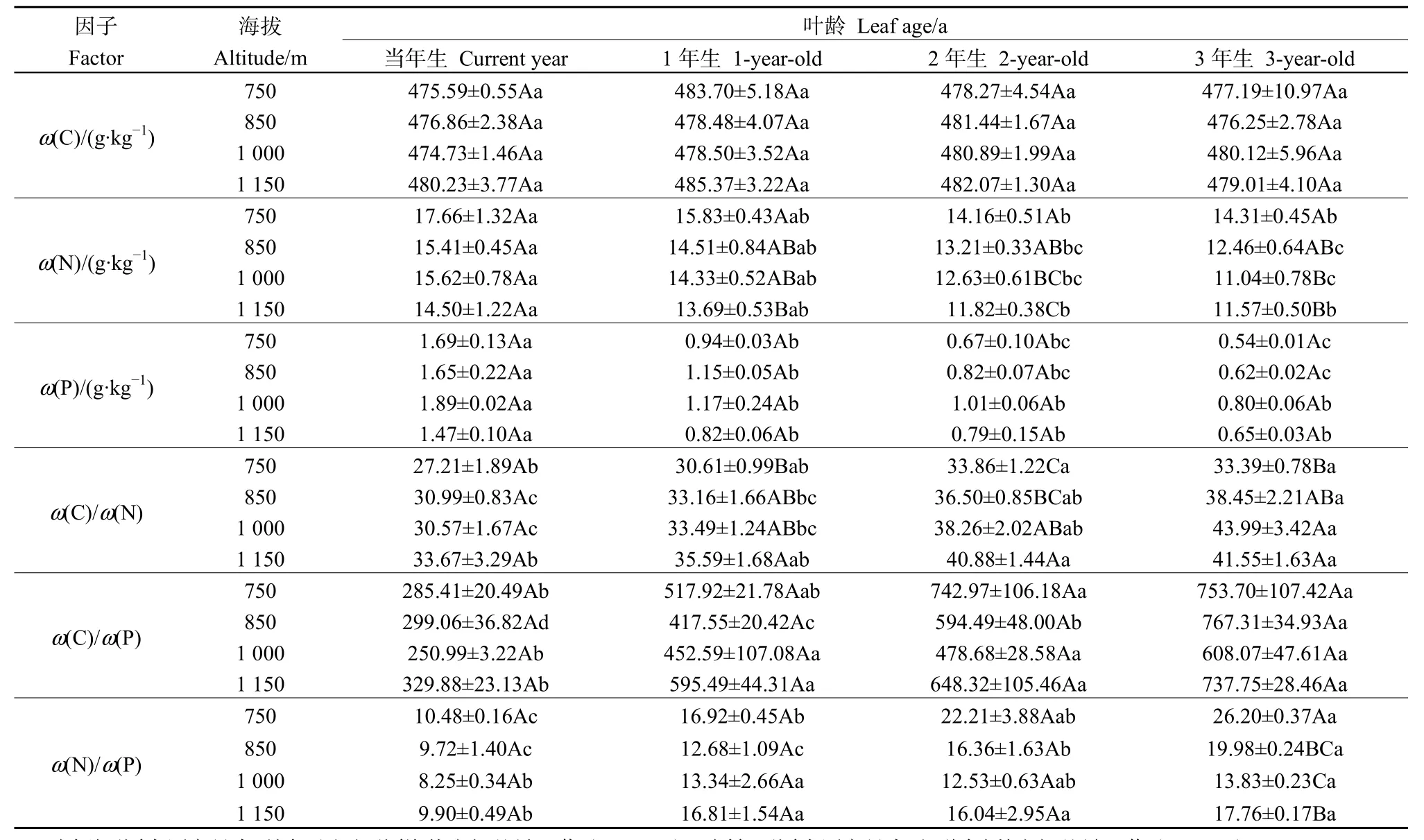
表4 不同海拔和叶龄杉木叶片碳氮磷含量及其化学计量比Table 4 Concentrations of C, N, P and their stoichiometric ratios of Cunninghamia lanceolata among different altitudes and foliar ages

表5 海拔、叶龄与杉木叶碳氮磷指标之间的Pearson相关系数Table 5 Pearson correlation coefficients between altitude, foliar age and C, N, P parameters of Cunninghamia lanceolata
3.3 海拔、叶龄与叶片因子之间的关系
海拔与 δ13C呈极显著的正相关关系,叶龄与δ15N之间呈显著的负相关关系,叶龄与P之间呈极显著的负相关关系,叶龄与 C/P、N/P之间呈极显著的正相关关系,海拔和叶龄都与N呈极显著的负相关关系,海拔和叶龄都与C/N呈极显著的正相关关系(表5)。说明海拔和叶龄对各元素有着显著的影响效果。在叶片因子中,只有C与δ13C含量之间呈极显著的正相关关系,其他因子与δ13C、δ15N之间均无显著的相关性(图2)。可以看出叶片营养元素并不是影响δ13C、δ15N含量变化的主导因素。
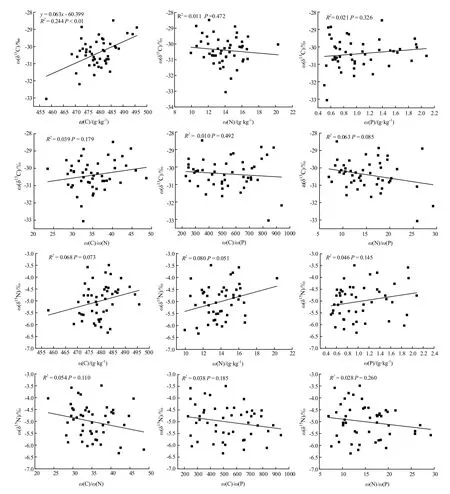
图2 杉木叶片养分与δ13C、δ15N之间的关系Fig. 2 Relationship between leaf nutrient and δ13C, δ15N of Cunninghamia lanceolata
4 讨论
4.1 海拔、叶龄对δ13C的影响
本研究中,海拔对杉木当年生叶片δ13C含量影响显著,随着海拔的升高,杉木叶片δ13C含量逐渐升高,说明杉木随着海拔的增加其叶片的WUE更高,并且当年生叶片对海拔的响应更明显。Oleary(1981)研究表明,C3植物的 δ13C的变化范围是-20‰--35‰,平均值为-26‰。杉木叶片δ13C的变化范围是-28.48‰--33.06‰,平均值是-30.39‰,在C3植物δ13C的变化范围之内。Li et al.(2017)利用已发表的文献以及来自中国东北样带和青藏高原的未发表数据,建立了中国C3陆生植物叶片δ13C的全国数据集,除了少部分亚乔木、落叶灌木和落叶乔木外(如落叶灌木和落叶乔木与海拔高度不相关),大多数C3植物的叶δ13C均受到海拔的影响,随着海拔的增加而增加。冯秋红等(2011)研究发现刺叶高山栎(Quercus spinosa)叶片δ13C含量会随着海拔的升高随之增加,认为是大气压强降低所引起的叶片扩散导度以及叶肉细胞导度降低导致δ13C含量增加。张鹏等(2010)在研究祁连山两种高山乔木时发现其叶片 δ13C含量也随着海拔的增加而增加,且其叶片δ13C含量与气候因子(年平均气温,年平均降水量)具有一定的相关关系,说明海拔变化引起的气候因子的改变,对高山乔木叶片δ13C含量起重要作用。海拔作为一个综合因子,其变化必然会导致气候因子中温度、降水、气压等的变化。就温度而言,随着海拔的增加,温度会逐渐降低,有部分研究表明,温度是影响植物叶片δ13C变化的主要因素之一(Hultine et al.,2000;旺罗等,2003),主要原因是温度的降低会导致叶片内部 CO2的扩散能力降低,从而减小了气孔的CO2导度,使叶片的δ13C增加。同时,温度的改变对植物生化酶活性的影响也是 δ13C变化的原因之一,低温使得生化酶活性降低,使其对13CO2的排斥能力减弱,从而导致δ13C增加(刘小宁等,2010)。海拔的变化同样会导致降水的变化,干旱区水分胁迫可诱导叶片气孔关闭,从而引起叶片内CO2浓度下降,使光合产物的δ13C值增大(张鹏等,2010),在低海拔地区,降水会随着海拔的增加而增加,叶片 δ13C有降低的趋势(张普等,2016;林晗等,2013),这与本研究区的结果相悖,可能是降水量超过阈值后降水量对植物叶片 δ13C的影响会逐渐减小(何春霞等,2010),这也说明研究区降水充足,植物生长未受到水分胁迫。此外,气压也影响植物叶片δ13C含量,但并不起主导作用,大气压随海拔升高而降低,CO2分压抑制光合作用导致的δ13C增加的同时 O2分压会抑制呼吸作用的进行,两者相互抵消,使得气压对植物δ13C的影响效果不明显(张普等,2016;刘小宁等,2010)。所以,影响本研究区杉木叶片 δ13C含量的气候因子可能主要是海拔变化导致的温度的改变。
叶龄对杉木叶片δ13C含量影响不显著,每个海拔梯度下不同叶龄的变化趋势都不尽相同,总体上随着叶龄的增加而降低,达到3年生时略有上升。孔令仑等(2017)发现亚热带杉木人工林不同叶龄之间的杉木WUE差异显著,随着叶龄的增加而降低,认为是不同叶龄之间的N、P养分的利用效率不同导致的。Werth et al.(2015)在研究两种红树林蕨类植物中发现 δ13C含量随着叶龄的增加略有下降,更容易在新叶中富集,这主要是植物在发育阶段会运输更多的养分到植物新叶中供新叶生长发育。本研究与前人们的研究大致相同,杉木不同叶龄之间δ13C含量差异不显著,这可能是不同叶龄之间C含量差异不显著有关,由于本研究中叶片采集时间是7月底,杉木当年生叶片基本上发育成熟,也说明研究区内杉木叶片在发育成熟之后其水分利用效率基本上没有很大的差异。
4.2 叶片养分对δ13C的影响
植物叶片δ13C与植物叶片养分密切相关,朴河春等(2004)在研究C3植物小果蔷薇(Rosa cymosa)时发现在海拔450 m的贵州茂兰至海拔1330 m的安顺一线,叶片δ13C含量随着海拔梯度的增加而增加,并发现叶片δ13C与N、P呈正相关,与C/N呈负相关,认为是植物叶片营养元素的相互作用导致δ13C发生这种变化。彭邵锋等(2016)在研究 11个山茶属(Camellia)植物叶片δ13C与养分关系时也发现叶片δ13C含量与C、N、P含量呈显著负相关,也进一步说明叶片δ13C含量可以指示叶片养分含量的高低。熊鑫等(2016)研究叶片δ13C与元素含量的关系时发现,鼎湖山优势树种叶片δ13C与叶片N含量呈极显著负相关,与叶片C/N呈极显著正相关。可以发现,在不同地区、不同树种之间,叶片δ13C与其养分含量不完全相同,这与不同树种之间的遗产特性和不同地区之间的气候条件不同有关。本研究区域属于杉木纯林,主要考虑气候条件的影响。本研究中,叶片δ13C与C含量呈正相关,叶片C含量反映植物的固碳能力,是植物光合呼吸作用的综合表现,说明杉木叶片δ13C可以反映杉木的固碳能力。植物叶片的N含量通过影响其细胞间CO2分压与大气CO2分压的比值,影响其δ13C含量,对于C3植物,大多数N通常存在于叶绿体、Rubisco以及与 C同化相关的其他结构中(Reich et al.,1995),因此,叶片N与羧化效率以及光合能力之间呈正相关,从而与植物叶片δ13C含量也存在正相关关系。本研究中,N含量与δ13C含量不存在显著的相关关系,且变化趋势相反。一般而言,随着海拔的升高,温度逐渐下降,根据生物地球化学假说,温度下降会使矿化速率降低,导致土壤可利用性N减少,最终植物叶片 N含量降低(Reich et al.,2004),温度可能是造成调查区杉木叶片N含量与δ13C含量没有显著相关性的主要原因。也有研究发现,一些植物的WUE与P含量呈正相关(Singh et al.,2010;Boogaard et al.,2010)。本研究中,随着海拔梯度的增加,杉木叶片P含量逐渐升高,在海拔1000-1150 m之间反而逐渐下降,生态系统中,土壤 P与植物 P存在耦合关系(杨思琪等,2017),而 N/P通常作为养分受限的判断指标(Koerselman et al.,1996),当N/P>16时,林分P限制;当N/P<14时,林分N限制;当N/P介于14和 16 之间,受到 N、P的共同限制(Xu et al.,2014),调查区海拔在750-1150 m之间,整个林分从P限制向N限制转变,但在1150 m处于N、P共同限制,在P限制条件下,δ13C较低,随着林分向N限制的转变,δ13C含量逐渐增加,说明P比N对δ13C含量更具有影响效果,而1150 m处出现的林分P限制可能导致叶片P含量对δ13C未产生显著影响,所以δ13C与叶片P含量的弱相关性可能可以反映林分的养分限制。
4.3 海拔、叶龄对δ15N的影响
本研究中,海拔对δ15N含量没有显著影响,随着海拔梯度的增加,叶片δ15N含量先降低,达到海拔1000-1150 m时开始增加。Zhou et al.(2016)研究青藏高原的高寒地区不同海拔梯度下植物叶片δ15N的变化时发现,温度和降水是导致叶片δ15N随着海拔升高而降低的主要原因,但青藏高原上还存在影响叶片δ15N的其他因素。Chen et al.(2015)在研究青藏高原高海拔样带栓皮栎(Quercus aquifolioides)叶δ13C和δ15N的分化格局时,发现青藏高原地区栓皮栎叶片 δ15N随着海拔梯度的增加而先增加后减少,这种变化趋势发生在海拔3300 m附近,并认为是土壤N素有效性变化导致的。Yi et al.(2006)研究发现高寒草甸植物δ15N随海拔升高会逐渐降低,但不显著,刘贤赵等(2018)在研究草本植物 δ15N海拔梯度变化时也发现植物 δ15N随海拔增加并未表现出明显的降低趋势,其主要原因可能是研究区域类海拔变化导致的温度和降水未能有效控制氮同位素的分馏。大部分研究表明,温度是通过影响土壤矿化、硝化和反硝化过程来调节 δ15N 的(Chang et al.,2017;Dawes et al.,2017)。降水增加导致植物 δ15N降低的原因是通过增加降水量导致植物通过叶片气孔或表皮直接吸收了大气降水中更多的 NH4+,而降水中的铵态氮的 δ15N往往较硝态氮的更少(Sutton et al.,1995)。在研究区的750-1000 m之间,温度降水可能参与调节了氮同位素的分馏,导致 δ15N随着海拔的增加而降低,但未产生差异,可以看出δ15N不及δ13C对海拔响应明显,δ13C比δ15N更能有效地指示气候环境的变化。但在1150 m δ15N的升高可能是土壤中N、P素共同调节导致的,因为δ15N与C/P和N/P的变化趋势一致,有研究表明,植物体内的 C/P、N/P一定程度上能反映土壤中P的供应状况(杨思琪等,2017;Koerselman et al.,1996)。研究区域内随着海拔的增加叶片由P限制向N限制转变,N限制导致植物获得的有效氮降低,δ15N含量随之降低,而在海拔1150 m处杉木叶片N/P介于14和16之间,又出现了N、P共同限制,林分N限制的减轻和P限制的加重提高了氮同位素的分馏,使得 1150 m δ15N含量高于750 m,土壤P限制进一步促进了氮同位素的分馏。
叶龄对叶片δ15N含量产生显著影响,随着叶龄的增加δ15N含量逐渐降低。葛露露等(2018)在研究滨海沙地不同树种人工林叶片时发现 δ15N均呈现出随叶龄增大而减小的趋势。本研究中N含量与δ15N虽然没有相关性,但变化趋势基本一致,δ15N在叶龄之间的差异可能是 N在不同叶龄之间的富集效应不同导致的。
4.4 叶片养分对δ15N的影响
关于叶片养分对δ15N的影响,大部分研究发现叶片养分对 δ15N的影响主要集中在 N对叶片δ15N 的影响(熊鑫等,2016;郑璐嘉等,2015;Hobbie et al.,2000),因为高浓度的N往往指示着高δ15N含量(Wang et al.,2019)。但本研究中,杉木叶片δ15N与N未产生显著相关关系。Hobbie et al.(2008)认为,叶片δ15N含量同叶片N含量之间的相关性反映了菌根对叶片 δ15N含量的影响,这说明研究区域内不同海拔梯度下杉木菌根效益差异可能不太明显。此外,其他叶片元素(C、P)对杉木叶片没有显著相关关系,可能与这些养分元素不是参与土壤矿化、硝化和反硝化的主要元素有关。
5 结论
通过对马鬃岭地区不同海拔梯度下不同林龄杉木人工林叶碳氮同位素以及 C、N、P及其化学计量比的研究得出以下结论,(1)导致马鬃岭地区杉木叶片 δ13C含量随海拔增加而增加的主要原因可能是温度改变杉木叶片气孔导度以及叶片生化酶活性导致,且当年生叶片比其他叶龄叶片对气候环境条件的改变更为敏感。(2)影响不同海拔梯度下杉木叶片 δ15N含量的主要原因可能是温度和降水以及土壤中N、P共同控制导致的。(3)马鬃岭地区杉木叶片养分元素对碳氮同位素的影响效应较小,碳氮同位素并不能有效反映植物的养分状况,但δ13C可能对环境的变化效应明显,δ15N可能对土壤养分的限制较为敏感。
