浙江省古田山自然保护区3种无尾两栖动物肥满度的比较
2019-11-04吴驰星泮琦斌
王 宇, 刘 盼, 吴驰星, 泮琦斌, 黎 慧, 邵 晨
(浙江师范大学 化学与生命科学学院,浙江 金华 321004)
0 引 言
动物的肥满度(body condition)是动物体通过取食和消化作用在其体内所积蓄的能量汇,它被认为是决定动物体适合度的一个重要因素[1].肥满度不仅影响到动物的存活率、生活史特征、动物与栖息环境的生态关系,以及动物自身的繁殖潜能;它还与动物体的健康状况和活力有关[2-4].利用动物的形态学参数、生物化学或生理学数据所构建的肥满度指数(condition index)不仅能很好地反映动物个体之间适合度的差异[5],还可以用于动物体能量或营养状况的评估[6];此外,肥满度指数还能在一定程度下反映出动物体内不同能量储存形式,如脂肪和蛋白质相对含量的多少[6].基于非伤害性的形态学方法所获取的动物体质量和体长数据所构建的肥满度指数计算方法[7],如比值指数法和残差指数法虽在实际应用中存在一定不足,但这些方法在某种程度上却能反映物种能量储存情况及生活环境质量情况[3],因此,在国内外得到大量的研究和应用.目前,动物的肥满度指数已广泛应用于生态学、生理学、生物化学、遗传学和养殖业等方面,所涉及的研究对象包括了鱼类[8]、两栖类[9]、爬行类[10]、鸟类[11]和哺乳类[12]等脊椎动物,以及无脊椎动物中的甲壳动物[13]和蛛形纲动物[14]等.
古田山国家级自然保护区地处浙江省西部的开化县境内,总面积约8 107 hm2,拥有丰富的动植物物种资源,是我国东部低海拔地区野生生物物种的天然贮存库和生物基因库.目前,以中亚热带常绿阔叶林为主的古田山自然保护区因其丰富的植物物种组成而建立了中国森林生物多样性固定样地,并可对样地进行永久性监测;此外,古田山保护区还是我国两栖爬行动物监测与研究的关键样区,诸多重要的科学问题将在这里开展长期的监测和研究[15].因而,深入了解古田山物种的营养状况,以及能量的摄取和储存情况是评估物种在该地区长期存活的重要依据,也是在该地区成功进行长期监测的重要影响因素.
调查显示,古田山有两栖动物2目7科17种,其中华南湍蛙(Amolopsricketti)、淡肩角蟾(Megophrysboettgeri)和花臭蛙(Odorranaschmackeri)是保护区内的优势两栖类物种[16].目前在有关两栖动物肥满度的研究中,尚未见到上述3个物种的相关报道.因此,本研究拟通过采用非伤害性的形态学研究方法构建上述3个物种不同的肥满度指数,以期为该类群物种在古田山自然保护区的长期监测和研究提供基础资料,同时为该地区两栖动物栖息环境的快速评估提供参考依据,并揭示物种在该地区的健康状况和营养状况.
1 材料和方法
1.1 样本采集及样本测量
2014年7—8月于夜间18点—次日凌晨2点在古田山国家级自然保护区的各山溪沟谷地带随机选择了26条1 000 m×6 m的样带对所遇见的华南湍蛙、淡肩角蟾和花臭蛙进行调查研究.用手持全球定位仪(GPS)记录在调查中所捕获的各样本的地理坐标,并依据《中国两栖动物及其分布彩色图鉴》[17]对各物种及个体进行识别鉴定.随后用数显游标卡尺(精确到0.1 mm)对各样本的头体长(即自吻端至泄殖腔孔的长度,SVL)进行测量,用电子天平(精确到0.1 g)称量各样本的体质量(body mass);并以是否出现婚垫来判定各样本的性别,以出现婚垫的雄性个体最小体长来判定各物种的雌性成体和亚成体[18].本研究中各物种亚成体的具体判定标准为:华南湍蛙个体的头体长小于37 mm,淡肩角蟾个体的头体长小于29 mm,花臭蛙个体的头体长小于34 mm.最后将各样本进行剪趾标记后原地释放.调查共记录到研究样本451只,其中华南湍蛙287只、淡肩角蟾53只、花臭蛙111只.
1.2 数据分析
分别用3种方法对古田山3种无尾两栖类的肥满度情况进行评估.首先使用Fulton的肥满度指数K[19],由于该方法仅简单考虑动物个体的体质量及体长的比值,因此,该方法自1908年从鱼类的自然种群研究及渔业管理中提出以来,在各大动物类群的肥满度研究中得到了极为广泛的应用;其次,根据Hayes等[20]的方法将BM和BL进行log处理后来进行评估,该方法对于数据具有较大误差或数据呈对数分布的物种应用较好,如在对老鼠肥满度的研究中,log法所获得的结果相较于其他17种肥满度指数具有最好的评估结果[21];最后使用SMI(scaled mass index)法来计算肥满度,SMI法是于2009年新提出的肥满度计算方法[2],该方法先基于体质量和体长之间固定的线性尺度关系将体质量进行标准化处理后,再计算物种的肥满度,虽然该方法并未在两栖动物肥满度的研究中与其他方法进行比较研究,但Maccracken等[22]认为在两栖动物的肥满度研究中SMI法能很好地反映物种真实肥满度情况.各指数的计算公式为:
Fulton肥满度指数K=M/L3;
Hayes肥满度指数log(BM)/log(BL)=lgM/lgL;
Peig肥满度指数SMI=Mi×(L0/Li)bSMA.
式中:M为各个体的体质量;L为各个体的体长;Mi为某物种第i个个体的体质量;L0为该物种的算术平均值;Li为该物种第i个个体的体长;bSMA是物种的尺度指数,是基于体质量M和体长L的标准主轴回归(standard major axis regression,SMA)估计得出的.
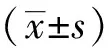
2 结 果
2.1 相同物种不同性别及不同年龄组肥满度的比较
不同性别的华南湍蛙,其肥满度指数在3种方法中均表现为雌性个体显著大于雄性个体;在不同年龄组中,K法显示华南湍蛙亚成体的肥满度指数显著大于成体,而在另外2种方法中则是成体的肥满度指数明显大于亚成体的肥满度指数(见表1).
肥满度指数的3种计算方法都表明淡肩角蟾中雌性个体高于雄性个体,且除K法外,另外2种指数均表现出明显差异;不同年龄组间,K法显示亚成体的肥满度指数显著高于成体,而另外2种方法则显示成体明显高于亚成体(见表1).
花臭蛙的肥满度指数在3种不同计算方法下均表现为雌性极显著的高于雄性;在除K法的另外2种方法中,花臭蛙在不同年龄组间的肥满度还表现出极显著的差异,且成体高于亚成体,而在K法的亚成体和成体之间,二者的肥满度指数没有明显的差异(见表1).

表1 不同性别、不同年龄组的3种无尾两栖动物肥满度变化及比较
注:*P<0.05表示有显著性差异,**P<0.01表示有极显著性差异.
2.2 不同物种的肥满度比较
用肥满度指数的3种不同计算方法比较花臭蛙、淡肩角蟾和华南湍蛙3个物种雄性个体、雌性个体、成体、亚成体及物种间的整体差异,结果显示,在雄性个体中3种方法均表现出显著的差异(见表2),进一步多重比较分析后提示,除K法的花臭蛙和淡肩角蟾雄性个体间的肥满度不具显著性差异外,其他的各组比较均表现出不同物种雄性个体间的显著肥满度差异.
此外,在不同物种的雌性个体、成体、亚成体及整体间的肥满度比较结果显示,上述各组比较应用K法并未能发现明显差异,而在log(BM)/log(BL)和SMI 2种方法下,上述各组比较均表现出极显著的差异(见表2),进一步的多重比较分析后显示,除花臭蛙和淡肩角蟾在log(BM)/log(BL)法中未表现出成体间及整体的差异外,其他的所有比较均有显著性的差异.

表2 不同物种相同性别、相同年龄组及整体的差异显著性比较
注:*P< 0.05表示有显著性差异,**P< 0.01表示有极显著性差异.
3 讨 论
动物的肥满度指数作为评估动物体适合度的重要指标,在一定程度上可反映动物体的能量储存状况或营养状况.一般认为动物体的肥满度指数越高,其能量储存状况或营养状况越好,体型也越胖或体质量更大[6];同时,具有较高肥满度指数的动物体在运动能力[23]、存活率[24]和繁殖潜力[25]上也显著优于肥满度指数较低的动物体;此外,动物体在具备更好生境质量和更多猎物数量的区域,其肥满度指数也将更高[26].然而,肥满度指数是一个与物种相关的经验性评估指数,目前除在少数类群存在肥满度标准以外(如在鳟鱼中,肥满度K=1表示该个体属于正常状态,K>1表示该个体营养状况较好,而K<1则表示该个体营养状况较差),大多数的动物类群均不具备相关的标准,也即在这些类群中没有一个确定性的标准来说明动物个体究竟是处于何种营养状况.因此,比较同一物种不同个体或不同物种间的肥满度对于揭示物种的营养状况和能量储存状况具有重要的作用.
3.1 不同性别之间肥满度的比较
3个物种的雌性个体的肥满度均大于雄性个体,这说明在这3个物种中,雌性个体的能量储存状况或营养状况均优于雄性个体.不同性别的两栖动物在生命活动中具有不同的能量分配模式:在非繁殖期,两性主要通过捕食大量的食物来育肥,其能量消耗主要用于捕食,所以此时两性之间的肥满度无明显差异,这与戴建洪等[27]在四川若尔盖湿地对中国林蛙的肥满度研究中得到的结论一致.在繁殖期,雄性个体除了将体内所储存的能量用于繁殖活动以外,还得花费大量的能量和时间用于寻找配偶、打斗和觅食等生命活动,而此时雌性个体为了能够增大繁殖的窝卵数,提高繁殖潜能,会将更多的能量用于繁殖活动,因此在繁殖期的繁殖活动前,雌性个体的能量储存状况将优于雄性个体;此外,由于在繁殖期内雄性个体的觅食时间相对于雌性也较少,因而其所能获得的能量或营养状况也将劣于雌性个体.因此,华南湍蛙、淡肩角蟾和花臭蛙在繁殖季节雌性个体的肥满度会显著高于雄性个体.金晨晨等[28]对黑斑侧褶蛙肥满度的相关研究及王寿兵等[29]对林蛙肥满度的研究结果均得到与此一致的结论;此外,在对鱼类[30]和哺乳类[31]等不同性别间肥满度的研究中,也发现具有雌性个体肥满度大于雄性个体的现象.
3.2 不同年龄之间肥满度的比较
不同年龄的动物用于维持生命活动所需的能量及对食物的摄取有明显的不同.同一物种不同年龄组的个体通常栖息于同一区域,然而由于成体体型较亚成体大,因而成体在食物竞争中占据较大的优势;其次,成体的头部相对亚成体较大,拥有较大头部的个体能够摄入猎物的体积也越大[32],因而成体较亚成体可获得更丰富的食物资源[33];再次,在同一空间内亚成体的密度远较成体大,亚成体较大的种群密度将导致个体之间的食物竞争加剧,并因此导致亚成体个体出现相对较差的营养状态[9];最后,成体和亚成体在能量分配上有所不同,成体主要将能量和营养物质储存在体内育肥以为繁殖活动服务,而亚成体并不储存能量而是将能量用于身体的生长.因此,华南湍蛙、淡肩角蟾和花臭蛙的成体肥满度要优于亚成体的肥满度.在其他相关研究中,虎纹蛙[9]、三疣梭子蟹[34]和布氏田鼠[12]等物种均出现成体肥满度优于亚成体的现象.
3.3 不同物种之间肥满度的比较
不同物种所占据栖息地的特征和质量是影响物种肥满度的重要因素[26].本研究中,华南湍蛙、淡肩角蟾和花臭蛙3个物种在古田山自然保护区的生态位分布并无明显的重叠,华南湍蛙主要分布于不同海拔高度水流湍急的区域,淡肩角蟾分布在高海拔水流较小的山溪附近,而花臭蛙则主要生活在低海拔水流较缓的地区[17-18].因此,在不同的栖息环境中,3个物种在食物资源的可获得性和可利用性上的差异是造成物种肥满度差异的重要原因;其次,人类的活动,如对栖息地的破坏和种植不同的农作物等,也会在一定程度上造成两栖动物栖息环境和食物资源的差异[35],并进而导致物种肥满度的不同;最后,动物自身对能量消耗的需求、能量转化效率,以及动物的运动能力、性成熟年龄、繁殖特点等诸多条件均可能对肥满度造成不同的影响,并最终导致不同物种间的肥满度不同.
3.4 不同肥满度指数之间的比较
肥满度指数自提出以来已有较大发展和变化,并先后出现比值指数法、残差指数法和体重尺度指数法等研究方法.比值指数法是通过应用动物的体长和体质量之间的比值来衡量个体的肥满度,如在本研究中所应用的Fulton的K法和log(BM)/log(BL)法.虽然比值指数法简单易操作,且其中的K法在渔业生产和管理中得到广泛应用,但由于该方法所获得的肥满度指数与体长L密切相关,且这种相关性在实际研究中不能被忽略,因此,物种的肥满度指数会受到体长差异的较大影响,尤其在进行物种间的肥满度比较时,其得到的肥满度指数往往很难反映该物种的真实状况,因此该方法的应用受到诸多的限制[20].其次,残差指数法是应用最小二乘法对体质量和体长之间的关系进行回归分析后用残差表示物种的肥满度指数.该方法在小型哺乳类和椋鸟类能很好地反映物种的能量储存情况.然而在实际研究中,物种的体长/体质量参数往往不能满足该方法所需要的较多前提假设(如:物种的体质量和体长功能关系必须线性、体长必须能精确反映物种的结构大小、体长不能与体质量有相关性等),且该方法极易导致Ⅰ和Ⅱ类错误.因此该方法的应用也受到较大的挑战[1].近年来发展起来的体重尺度指数法是基于体质量为中心尺度的原则所建立的肥满度计算方法,已有研究发现该方法所得到的肥满度指数与物种的能量储存具有非常显著的相关性,且对于不同物种和异速生长物种之间的肥满度比较,或同一物种不同性别和不同年龄之间的比较具有重要的应用价值[2-3].此外,本研究结果也揭示了关于同一物种不同性别及不同物种间的比较,SMI法相对于其他2种方法肥满度指数更为敏感.
