高脂日粮对肥胖易感和肥胖抵抗小鼠骨性能的影响以及槲皮素的干预作用
2019-10-30周银芝夏淑芳段晓梅施用晖乐国伟
周银芝,夏淑芳,段晓梅,颜 彪,施用晖,2,乐国伟*,2
(1.江南大学 食品学院,江苏 无锡 214122;2.食品科学与技术国家重点实验室,江南大学,江苏 无锡 214122)
骨质疏松症和肥胖这两种身体组成紊乱的疾病在全球范围内正以高比例增长[1]。但是关于肥胖对骨骼的影响还存在争议。机械负荷可以通过减少细胞凋亡促进成骨细胞和破骨细胞分化来促进骨形成[2]。由高体重产生的机械负荷使得人们普遍认为肥胖可以预防骨丢失和骨质疏松[3]。然而Compston等人发现,肥胖不仅不会保护更年期妇女不得骨质疏松症,反而会增加她们股骨骨折的风险[4]。经骨密度校正后,较高的体重是所有骨质疏松性骨折的危险因素[5]。
肥胖患者有较高的完全代谢率,机体的氧化应激程度增加[6]。肥胖个体通过抗氧化酶水平降低表现出抗氧化防御系统降低[7],降低体重对于减轻氧化应激是十分重要的[8]。高脂日粮刺激下,伴随着肥胖发生,小鼠股骨发生氧化应激并诱发骨代谢失衡[9]。具有抗氧化功能的功能因子可以有效缓解氧化应激。硫辛酸可通过清除自由基、调整氧化还原状态,改善小鼠肝脏脂代谢功能[10],槲皮素可有效提高机体抗氧化能力,缓解高脂膳食造成的氧化应激,改善血脂代谢[11]。高脂膳食下的肥胖抵抗、中等体重、肥胖易感三种肥胖表型小鼠血液CAT、TAOC均比正常膳食小鼠显著下降,MDA含量显著增加,血浆低密度脂蛋白含量也呈显著上升的趋势,而肥胖抵抗小鼠氧化应激程度比肥胖小鼠相对减轻[12]。因此将肥胖抵抗和肥胖易感小鼠分开研究更能够说明高脂日粮对小鼠股骨氧化还原状态的影响。
作者通过饲喂高脂日粮诱导出肥胖易感和肥胖抵抗两种表型,探究短期和长期饲喂高脂日粮的OP和OR小鼠的骨性能和股骨氧化还原状态的变化。通过测定相关基因的表达进一步探究不同周期高脂日粮影响OP和OR小鼠骨性能的可能作用机制以及抗氧化功能因子槲皮素对此的干预作用,为肥胖抵抗和肥胖易感人群预防和干预骨质疏松提供理论基础。
1 材料与方法
1.1 实验试剂及仪器
槲皮素:纯度为99%,美国sigma公司;牛血清蛋白:上海生工物程有限公司;氧化性谷胱甘肽(oxidized glutathione,GSSG),还原性谷胱甘肽(glutathione,GSH)标准品:中国医药集团化学试剂有限公司;丙二醛(MDA)、总抗氧化能力(T-AOC)试剂盒:南京建成生物科技有限公司;荧光染料SBY:Bioneer公司 ;DEPC:Generay Biotech 公司 ;Trizol裂解液:美国Biomiga公司;反转录各种酶和缓冲液以及dNTPs:大连宝生物工程有限公司;基因引物:Generay Biotech公司;其余试剂均为分析纯。In-Vivo DXS PRO全自动高分辨X光机:德国Bruker公司;手持式组织匀浆器PT1200:Kinematica公司;SpectraMax M5/M5e酶标仪:美国Molecular Devices公司;恒温水浴锅:上海精宏实验设备有限公司;XB70制冰机:美国GRANT公司;Forma超低温冷冻冰箱:美国Thermo公司;AB204-N分析天平:梅特勒-托利多公司;STARTER 3100实验室pH计:上海奥豪斯仪器有限公司。
1.2 实验动物和喂养
110只3周龄C57/BL6雄性小鼠,购自南京大学模式动物研究所,随机分为饲喂正常日粮的对照组(n=20),高脂日粮组(n=90),自由采食饮水,饲养温度 (23±2)℃,湿度60%,12/12小时昼夜循环光照。7 w后,将饲喂高脂日粮的小鼠按体重从低到高排序,体重最低的1/3称为肥胖抵抗(obesity resistant,OR,n=10),体重最高的 1/3 的称为肥胖易感(obesity prone,OP,n=10),处死。 第二批,高脂饲喂7 w分出OR和OP后,给肥胖和抵抗小鼠的高脂日粮中均添加0.4%槲皮素得到肥胖易感+槲皮素组(OP+Q,n=10)和肥胖抵抗+槲皮素组(OR+Q,n=10),只喂高脂日粮不添加槲皮素的称为肥胖抵抗高脂对照组(OR,n=10),肥胖易感高脂对照组(OP,n=10),17周末处死小鼠。饲料配方见表1。
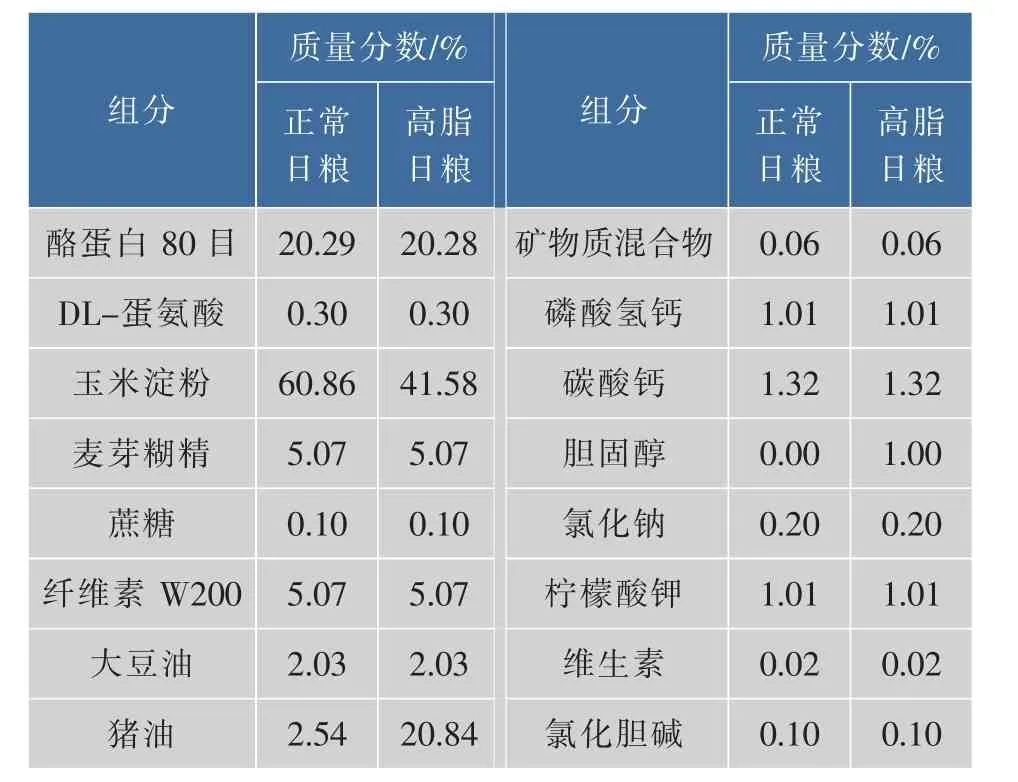
表1 饲料配方Table 1 Composition of diets
1.3 实验方法
1.3.1 组织样品制备各组小鼠禁食12 h,乙醚麻醉后断颈法处死小鼠,在冰浴上迅速取出股骨和胫骨;用生理盐水中浸泡过的纱布将左腿股骨擦除多余的附着组织后,取部分添加0.9%的预冷生理盐水制备10%的匀浆液,置于-20℃备用;剩余左腿股骨至于Trizol中用于RNA提取;其余样品液氮冷冻后置于-80℃超低温冰箱保存。
1.3.2 氧化还原状态指标测定MDA、T-AOC测定均按照南京建成生物科技有限公司试剂盒说明书操作。GSH和GSSG含量用荧光分光光度法测定。
1.3.3 总RNA提取及qRT-PCR用生理盐水中浸泡过的纱布擦除股骨上附着的组织后至于1 mL Trizol中,用灭酶剪刀将股骨剪碎,再用灭过酶的电动匀浆器冰浴匀浆。匀浆后放置15 min后,10 000 r/min离心15 min。吸取离心后的上清液加入200 μL氯仿混匀,静置15 min后,10 000 r/min离心15 min。吸取上层水相400 μL,加入等体积的-20℃预冷的异丙醇和等体积预冷盐溶液,混匀4℃沉淀过夜。其余操作按照Trizol法提取股骨总RNA,反转录得到 cDNA。实时荧光定量 PCR(quantitative Real-time PCR)检测小鼠股骨mRNA水平,引物序列见表2。

表2 引物序列Table 2 Sequences of the primers
1.3.4 骨性能参数测定在生理盐水中浸泡过的纱布擦除右腿股骨附着的组织后,测定以下指标。
骨密度:用In-Vivo DXS PRO全自动高分辨X光机测定股骨密度;骨长度(length,mm):用游标卡尺测定股骨头的近侧端至远侧末端;骨径:用游标卡尺测定骨干中点;骨强度(N):采用质构仪进行三点弯曲试验,将样品置于支撑架上,固定两端,跨距6 mm,探头加载速度10 mm/s直至股骨断裂获得最大载荷值;骨干质量 (dry weight,mg):将股骨于105℃常压烘干至恒质量;灰分重(mg):将股骨置于坩埚中,在电炉上碳化至不再产生黑烟后,放于恒质量后的坩埚中在马弗炉中550℃灰化至恒质量,计算灰分含量;股骨钙质量分数(mg/g dry weight):EDTA滴定法测定钙含量;股骨磷质量分数(mg/g dry weight):钒钼酸铵比色法。
2 结果与分析
2.1 高脂日粮不同周期肥胖易感和肥胖抵抗小鼠骨性能的变化
2.1.1 短期高脂日粮对OR和OP小鼠骨性能的影响由表3可知,饲喂高脂日粮7 w时OP组小鼠的体质量显著高于C组和OR组(P<0.05),并且达到了C组小鼠体质量的1.2倍以上,出现了肥胖易感和肥胖抵抗两种表型。
OR组小鼠股骨矿物质密度、生物力学性能指标骨强度、直径、灰分质量、干质量、股骨钙质量分数均显著低于C组(P<0.05),骨长度与C组相比无显著性差异。提示短期饲喂高脂日粮会使降低OR小鼠的骨性能。
OP组小鼠体质量显著高于C组小鼠,其股骨灰分质量、骨干质量显著高于C组小鼠(P<0.05),其矿物质密度、骨强度、股骨钙含量、骨长度分别比C 组高出 11.7%、3.1%、1.6%、13.4%(P>0.05), 提示高脂日粮会使OP小鼠的骨性能升高。有研究表明,短期高脂日粮喂养会使生长期小鼠的骨性能增加,骨量变大,但是经体重校正后的骨密度和骨强度都呈下降趋势[13],骨量较大并不意味着骨性能更好[2],肥胖小鼠骨性能增加“可能是为了支持过重体质量的代偿反应”[13]。OP组小鼠的骨密度和骨强度经体质量校正后的骨密度/体质量(BMD/Weight)、骨强度/体质量 (Strength/Weight)值低于OR组和C组(P>0.05)。提示OP小鼠虽然骨量增加但是骨性能有下降的趋势,结果见表3。
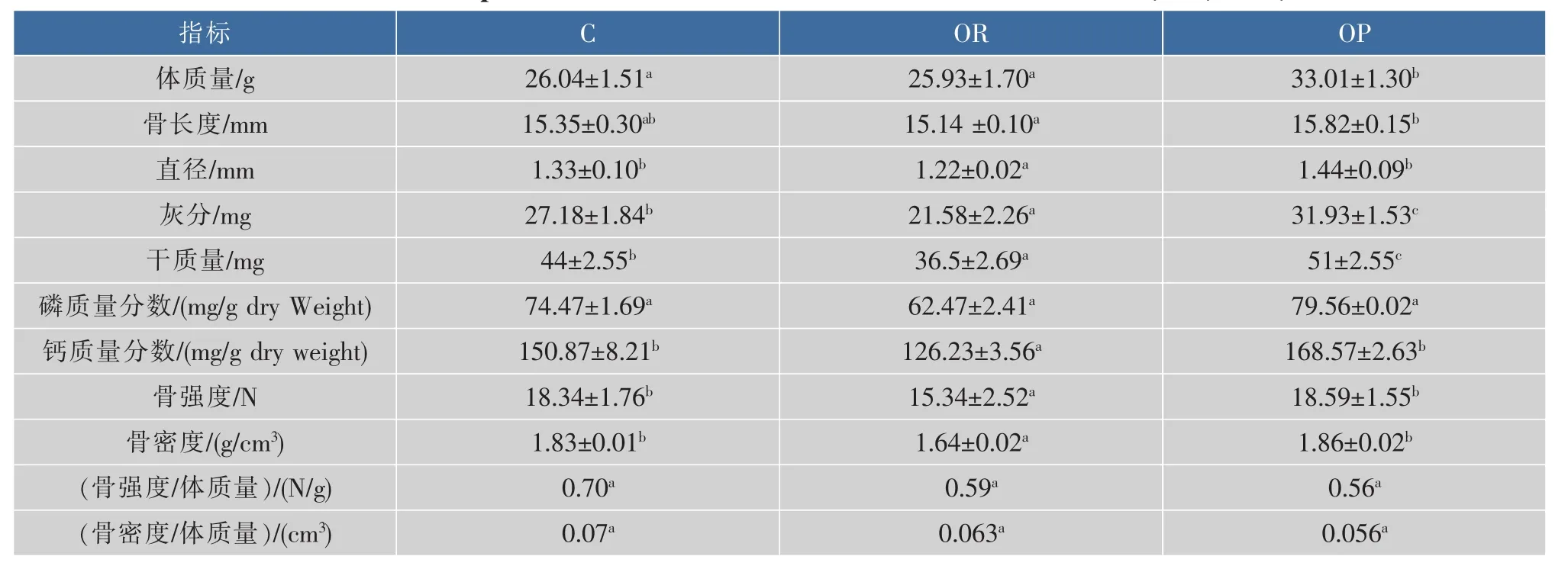
表3 短期高脂日粮肥胖易感和肥胖抵抗小鼠的骨性能(x±s,n=10)Table 3 Femur parameters of OR and OP mice after short term HFD(x±s,n=10)
2.1.2 长期高脂日粮对OR和OP小鼠骨性能的影响由表4可知,饲喂高脂日粮17周时,OR和OP小鼠的体重均显著高于C组小鼠(P<0.05),说明OR小鼠对肥胖的抵抗能力是有限的,长期饲喂高脂日粮OR小鼠的体质量会显著升高。
饲喂高脂日粮17周时,OR组小鼠的骨矿物质密度、生物力学性能指标骨强度、直径、灰分质量、干质量、股骨钙质量分数都显著低于C组 (P<0.05),提示长期饲喂高脂日粮会影响OR小鼠股骨的生长,降低OR小鼠的骨性能。
饲喂高脂日粮17周时,OP组小鼠的股骨骨长度和直径与C组小鼠相比无显著差异(P>0.05)。但是其股骨的钙质量分数、骨强度、骨密度均低于C组小鼠(P<0.05),提示长期饲喂高脂日粮,对OP小鼠股骨的生长无显著性影响,但是会使OP小鼠的股骨性能降低,经体重校正后,OP组小鼠的骨强度,骨密度均显著低于OR组和C组小鼠。
添加0.4%的槲皮素,OR+Q组小鼠的体质量相对OR小鼠没有显著性变化,其股骨矿物质密度、骨强度、灰分质量均恢复到C组水平;OP+Q小鼠的体质量显著低于OP小鼠,仍显著高于C组小鼠,OP+Q组小鼠股骨矿物质密度、骨强度、干质量、股骨钙质量分数均得到恢复,见表4。
2.2 高脂日粮不同周期肥胖易感和肥胖抵抗小鼠股骨氧化还原状态的变化
如表5所示,和C组相比,OP组和OR组小鼠股骨抗氧化能力指标T-AOC和GSH/GSSG均水平显著降低(P<0.05),脂质过氧化物标志物MDA水平显著升高(P<0.05),表明OP和OR小鼠股骨抗氧化能力降低,氧化还原状态失衡。OR小鼠抗氧化指标均高于OP小鼠 (P>0.05),MDA水平高于OP小鼠(P>0.05),说明OP小鼠抗氧化应激的能力比OR小鼠低,此结果与OP组小鼠的骨密度/体质量、骨强度/体质量低于C组和OR组小鼠一致。

表4 长期高脂日粮周肥胖易感和肥胖抵抗小鼠的股骨性能(x±s,n=10)Table 4 Femur parameters of OR and OP miceby long term HFD(x±s,n=10)

表5 短期高脂日粮股骨氧化还原状态(x±s,n=10)Table 5 Femur redox statebyshort term HFD(x±s,n=10)
17周时,和C组相比,OR组小鼠和OP组小鼠抗氧化能力指标T-AOC和GSH/GSSG水平均显著降低(P<0.05),脂质过氧化物标志物MDA水平均显著升高(P<0.05),说明长期高脂日粮破坏OR和OP小鼠股骨抗氧化应激的能力。添加0.4%槲皮素后,OR和OP组小鼠股骨氧化还原状态均得到恢复,见表6。

表6 长期高脂日粮股骨氧化还原状态(x±s,n=10)Table 6 Femur redox statebylong term HFD(x±s,n=10)
2.3 高脂日粮不同周期肥胖易感和肥胖抵抗小鼠成骨和破骨相关基因表达
骨骼是一个不停更新的活跃器官,骨骼的更新是一个包含破骨细胞骨吸收,成骨细胞骨形成的过程[14]。所以骨量可以在任何时期反映骨形成和骨吸收之间的平衡[15]。Runx2是一种转录激活因子,是成骨细胞分化所必需的[16],能促进间充质干细胞的成骨分化过程并调节成骨细胞的成熟,Runx2基因敲除小鼠成骨细胞无法分化,软骨无法矿化[17]。Bglap2是成骨细胞成熟的标志物,监测它的表达可以监测成骨细胞的分化[18]。RANKL正常情况下由成骨细胞合成,与破骨细胞及其前体表达的RANK结合,促进破骨细胞前体向破骨细胞分化并抑制破骨细胞凋亡,造成骨吸收增加。OPG由成骨细胞分泌,其作为RANKL的诱骗受体,与RANKL结合后使后者不能发挥作用,OPG是骨吸收强抑制剂,RANKL的天然拮抗剂[19]。RANKL/OPG上升破骨细胞的数量增加活性增强。组织蛋白酶K(cathepsin K,CTSK)是破骨细胞重要的功能分子,参与降解骨有机基质,并可以通过调控破骨细胞凋亡控制破骨细胞数量,高脂膳食小鼠中骨吸收和破骨细胞分化增强可能与CTSK表达上调有关。高脂日粮造成骨组织氧化应激,骨组织表现出低的抗氧化能力、氧化还原状态向氧化态改变、ROS水平升高[20]。抗氧化功能因子槲皮素能够效提高机体抗氧化能力,缓解高脂日粮造成的氧化应激,改善血脂代谢[11]。核因子E2相关因子 2(nuclear factor E2-related factor 2,Nrf2)是调节抗氧化应激反应的重要转录因子,在机体氧化还原状态稳态的维持中起关键作用。Nrf2能够通过清除ROS维持骨重建平衡[21]。醌氧化还原酶(NADPH quinine oxidoreductase,NQO1)是Nrf2的下游抗氧化酶基因,NQO1能够增强饲喂高脂日粮小鼠骨组织的抗氧化能力[22]。
2.3.1 7 w肥胖易感和肥胖抵抗小鼠成骨和破骨相关基因表达如图1所示,与C组相比,高脂日粮7 w时,OR组小鼠Bglap2、Runx2的表达无显著变化,RANGKL/OPG、CTSK 的表达均显著上调(P<0.05),而 Nrf2、NQO1 的表达显著上调(P<0.05)。 提示短期饲喂高脂日粮能够上调OR小鼠破骨细胞形成相关基因的表达,打破骨重建平衡,从而降低OR小鼠的骨性能。OR小鼠股骨Nrf2被激活,上调下游基因NQO1的表达来提高股骨的抗氧化能力。
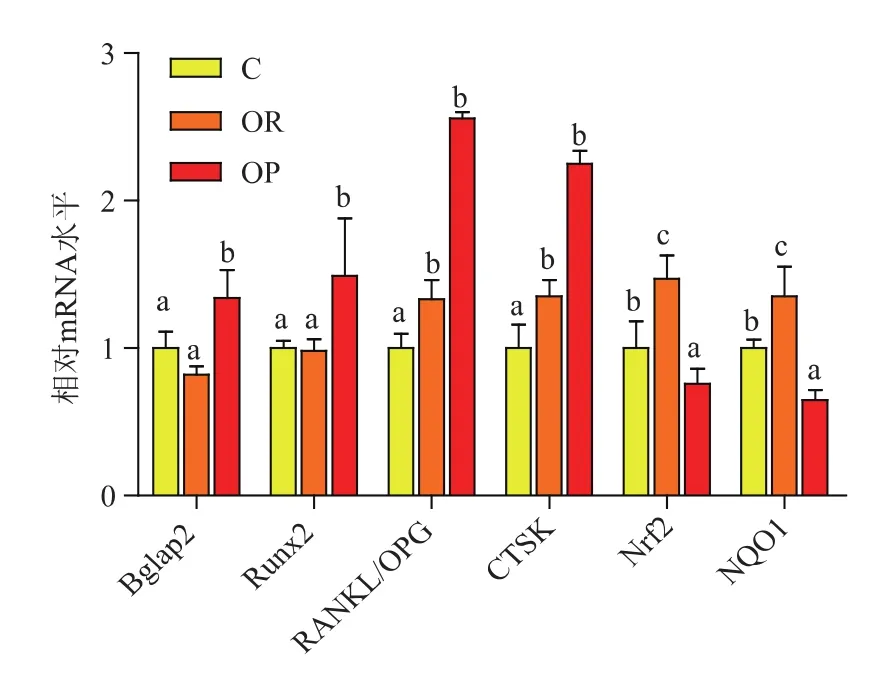
图1 短期高脂日粮OR和OP小鼠相关基因的表达Fig.1 Gene expressions of OR and OP mice after short term HFD
与C组相比,OP组小鼠Runx2、Bglap2的表达分别显著上调 1.34和 1.49倍 (P<0.05),RANGKL/OPG、CTSK的表达分别上调 2.56和 2.25倍 (P<0.05),提示短期饲喂高脂日粮,OP小鼠可能通过增强成骨提高骨性能,破骨细胞分化水平高于成骨细胞分化水平导致骨性能有下降的趋势,与骨性能的结果具有一致性。Nrf2、NQO1的表达均显著下调(P<0.05),提示短期饲喂高脂日粮OP小鼠股骨已经出现严重氧化应激。Nrf2、NQO1下调可能导致破骨基因RANGKL/OPG,CTSK的表达上调,导致骨性能降低。
2.3.2 17 w肥胖易感和肥胖抵抗小鼠成骨和破骨相关基因表达如图2-3所示,高脂日粮17 w时,与C组相比,OR组小鼠Bglap2、Runx2的表达显著下调(P<0.05),RANGKL/OPG、CTSK 的表达显著上调 (P<0.05), 而 Nrf2、NQO1的表达显著下调 (P<0.05)。提示长期高脂日粮能够通过增强破骨细胞分化相关基因的表达,抑制成骨细胞分化相关基因和氧化还原相关基因的表达降低肥胖抵抗小鼠骨性能。
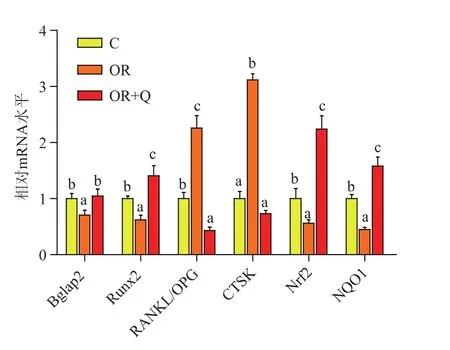
图2 长期高脂日粮OR小鼠相关基因的表达Fig.2 Gene expressions of OR mice after long term HFD
添加0.04%的槲皮素后,与C组相比,OR+Q组小鼠 Runx2的表达显著上调 (P<0.05),RANGKL/OPG、CTSK 的表达显著下调 (P<0.05),Nrf2、NQO1的表达显著上调(P<0.05)。提示槲皮素能通过增强骨骼的抗氧化能力,提高成骨细胞的分化,抑制破骨细胞的分化来改善肥胖抵抗小鼠的骨性能。
与C组相比,OP组小鼠Bglap2、Runx2的表达显著下调(P<0.05),RANGKL/OPG、CTSK 的表达显著下调 (P<0.05),而Nrf2、NQO1的表达显著下调(P<0.05)。提示长期饲喂高脂日粮能够通过促进破骨细胞形成相关基因的表达、抑制成骨细胞形成相关基因和氧化还原相关基因的表达影响肥胖易感小鼠的骨性能。

图3 短期高脂日粮OR小鼠相关基因的表达Fig.3 Gene expressions of OP mice after long term HFD
与C组相比,OP+Q组小鼠Bglap2、Runx2的表达显著上调(P<0.05),RANGKL/OPG、CTSK 的表达显著下调 (P<0.05),Nrf2、NQO1 的表达显著上调(P<0.05)。提示槲皮素能通过增强肥胖易感小鼠股骨的抗氧化能力,提高成骨细胞的分化,降低破骨细胞的分化来改善肥胖易感小鼠的骨性能。
3 讨 论
高脂日粮喂养时,OR小鼠采食量显著低于OP小鼠,但是OR小鼠的营养物质消化率显著高于OP小鼠[11],因此OR小鼠的体重较轻并不是因为营养物质摄入不足导致的,OR小鼠可能通过提高营养物质的消化率、控制采食量来控制体质量,本实验研究发现OR小鼠这种能力是有限的,长期饲喂高脂日粮体质量也会增加。
本研究发现,短期饲喂高脂日粮的OR小鼠股骨抗氧化能力下降,破骨细胞分化水平升高,股骨骨径变细,骨矿物质含量降低,骨密度、骨强度下降,骨性能降低,OR小鼠通过上调Nrf2、NQO1的表达来增强股骨的抗氧化能力。OP小鼠骨灰分含量和骨干质量显著高于C组小鼠,其骨长度、钙含量、骨强度、骨密度分别比C组高出3.1%、11.7%、13.4%、1.6%,但是经体质量校正的骨密度(BMD/Weight)、骨强度(Strength/Weight)均低于 C 组和 OR组(P>0.05),虽然没有显著统计学差异,但是呈现了低于C组和OR组的趋势。基因结果显示,OP小鼠成骨细胞分化水平升高,但是破骨细胞分化水平高于成骨细胞分化水平,Nrf2、NQO1的表达下调,TAOC和GSH/GSSG水平显著降低(P<0.05),MDA水平显著升高(P<0.05),股骨氧化还原状态失衡,引起破骨细胞分化增强。Wang Luan等人的研究表明,肥胖对生长旺盛阶段的幼鼠股骨骨量的增加不能充分代偿体质量的增加[13],由此推测OP小鼠可能通过增加成骨细胞分化来支撑过质量的体重带来的机械负荷,但是股骨出现氧化应激引起破骨细胞分化增强,且高于成骨分化水平,所以OP小鼠校正后的骨性能有降低的趋势。白静[11]等人的研究发现,高脂日粮喂养的OR小鼠机体的抗氧化能力强于OP小鼠,这与我们短期高脂日粮的研究结果是一致的。饲喂短期高脂日粮时,OP小鼠的股骨抗氧化能力显著低于OR组小鼠,解释了OP小鼠经体重校正过的骨强度和骨密度均呈现低于C组和OR组小鼠的趋势。
长期饲喂高脂日粮的OR和OP小鼠的骨性能各指标均显著降低,成骨细胞分化水平均显著降低,破骨细胞分化水平均显著升高,股骨均出现氧化应激,骨性能均降低。肥胖抵抗小鼠早期有着更强的抵御氧化应激的能力,可被长期高脂饮食破坏[23],饲喂高脂日粮17 w时,OR小鼠股骨的抗氧化指标降低,MDA水平显著升高,且与OP小鼠无显著差异,OR小鼠长期饲喂高脂日粮后股骨的抗氧化能力也被破坏。越来越多的研究表明,具有抗氧化功能的功能因子可以改善高脂日粮引起的氧化应激[10,24],槲皮素可以通过减轻股骨的氧化应激,增加成骨细胞的分化,抑制破骨细胞的分化来提高骨性能,因此加入槲皮素进行干预后,OR和OP小鼠的氧化还原状态和骨性能均得到恢复,槲皮素的作用机制还需进一步探究。
4 结语
综上所述,高脂日粮会破坏肥胖易感和肥胖抵抗小鼠的股骨氧化还原状态,引起氧化应激,饲喂短期高脂日粮OR小鼠抵御氧化应激的能力强于OP小鼠,OR小鼠骨性能呈现强于OP小鼠的趋势。长期高脂日粮破坏了OR和OP小鼠抵御氧化应激的能力,两种肥胖表型小鼠的骨性能均显著降低。氧化应激可能通过影响成骨细胞分化和破骨细胞分化相关基因的表达,降低骨性能,添加槲皮素能通过减轻两种小鼠的氧化应激提高骨性能。槲皮素在改善高脂饮食引起的骨质疏松方面具有潜在的应用前景。
